

Курс_лексия_Сабурова-2
.pdf
|
|
|
С18 |
|
|
С18 |
|
22АТФ |
||||||||
|
|
|
|
|
|
|
-1 |
|
|
|
|
-1 |
|
|||
|
|
|
|
2 |
|
|
2 |
|
|
|
||||||
|
|
|
8раз |
|
|
|
|
|
|
|
|
|||||
7раз |
*5=40АТФ- |
8раз |
|
|
||||||||||||
*5 |
|
|
2=38АТФ |
*5 |
|
|
|
|
||||||||
|
|
|
|
|
|
|||||||||||
35АТФ |
|
|
|
|
|
|
|
|
40АТФ |
|
||||||
|
|
|
=9 |
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
=9 |
|
|
|||
|
|
|
26 |
|
|
|
|
|
ЦТК – |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
12АТФ 12 26 = |
|
|
|
|
|
|
|||||||
|
|
|
312АТФ |
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
35+38+40=113 113 + 312 +22 = 447-4(активация жирных кислот и глицерина) = 443 АТФ.
Суммарное уравнение β – окисления, напр. пальм. к-ты может быть представлено таким образом.
cosКОА + 7ФАД+7 НАД +7 HSКОА |
|
|
|
8 |
+ 7 ФАД +7 |
|
|
|
НАД Во многих тканях β – окисление - важный источник энергии. Это ткани с
высокой активностью ферментов ЦТК и дыхательных цепей скелетных мышц, сердечной мышцы, почек. Эритроциты, в которых отсутствуют митохондрии, не могут окислять жирные кислоты. Жирные кислоты не служат источником энергии для мозга и др. нервных тканей, т.к. они не проходят через гематоэнцефалический барьер. При голодании, длительной физической работе и в случаях, когда клетки не получают достаточного количества глюкозы, жирные кислоты используются многими тканями, как основной источник энергии. В отличие от других тканей мозг и нервные ткани не используют жирные кислоты как источник энергии. В печени часть жирных кислот превращается в кетоновые тела, которые окисляются мозгом, нервной тканью, мышцами, обеспечивая достаточное количество энергии для синтеза АТФ, и уменьшая потребление глюкозы.
Литература:
А. Основные: 1. Биохимия. Т.Т.Берѐзов, Б.Ф.Коровкин. 2010 2. Биохимия. Б.А.Строев. 1986
Б. Дополнительные:
1. Медицинская биохимия. С.М.Рапапорта. 1976
11

2.Биохимия. Р.Страер. 1985
3.Северин. 2003
Лекция № 3 Обмен фосфолипидов
План:
1.Синтез триглицеридов и фосфолипидов.
2.Синтез высшых жирных кислот.
3.Метаболизм Ацетил КоА.
Метаболизм фосфолипидов тесно связан со многими процессами в организме, образованием и разрушением мембранных структур клеток, формированием липопротеидов , желчи, образованием в альвеолах легких поверх ностного слоя, предотвращающего слипание альвеол во время вдоха. Нарушение обмена фосфолипидов – причина многих заболеваний, в частности респираторного дистресссиндрома новорожденных, жирового гепатоза.
Переваривание фосфолипидов происходит в duodenum под действием ферментов фосфолипазы. Источники фосфолипидов: желток яйца, сливочное масло, печень.
В переваривание фосфолипидов участвуют ферменты фосфолипазы
А1, А2, Д, С.
Фосфолипаза А1 - отщепляет R1СООН, А2 – отщепляет ненасыщенную жирную кислоту. Фосфолипаза, А2 содержится в яде змей и скарпионов и при их укусе происходит гемолиз эритроцитов. В кишечнике эти две фосфолипазы действуют одновременно.
Образовавшиеся жирные кислоты в стенке тонкого кишка всасываются с участием желчных кислот. Фосфорная кислота всасывается в виде натриевых и калиевых солей. Азотистые основания всасываются в виде своих активных соединений.
Холин+АТФ+ЦТФ ЦДФ-холин – В таком виде всасывается, затем распадается и остается холин. В кишечной стенке происходит ресинтез фофосфолипидов.
Для синтеза фосфолипидов необходимы глицерин высшие жирные кислоты,Н3РО4 и азотистые основания.
Ресинтезированные фосфолипиды транспортируются в составе хиломикронов по лимфатической системе, поступают в верхний грудной проток и поступают в печень.
Синтез высших жирных кислот
12

Синтез высших жирных кислот протекает в цитоплазме.Основным компонентом является малоновая кислота, малонил КОА. Он образуется
путем присоединения к ацетил-КоА углеродного атома в виде  . 1)Сущность этой реакции состоит в том, что малонил-КоА находится на более высоком энергетическом уровне, чем ацетил-КОА и легче вступает в процесс синтеза.
. 1)Сущность этой реакции состоит в том, что малонил-КоА находится на более высоком энергетическом уровне, чем ацетил-КОА и легче вступает в процесс синтеза.
2)В реакциях участвует целый комплекс ферментов мультэнзимсинтетаза высшей жирной кислоты, в состав которой входит 7 ферментов.
Каждый из ферментов, входящей в этот комплекс, представляет собой белок, который называется ацилпереносящий белок – АПБ. Простетической группой является фосфопантотеин, единственной SH –группе принадлежит особая роль в синтезе жирных кислот.
3)В реакциях синтеза участвует НАДФ , источником которого является пентозный цикл окисления глюкозы.
, источником которого является пентозный цикл окисления глюкозы.
Впроцессе синтеза, ацетилКОА вступает в реакцию с ацилпереносящим белком (АПБ), образуется ацетилацилпереносящий белок.
.
13
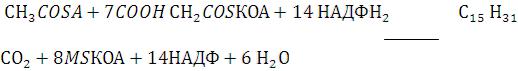
Дальше цикл реакции повторяется. Допустим, что при синтезе
польметиновой к-ты ( ), образованием бутирила завершается лишь первый из семи циклов, в каждом из которых началом является присоединение молекулы малонил АПБ. к карбоксильному концу растущей цепи жирной кислоты.
), образованием бутирила завершается лишь первый из семи циклов, в каждом из которых началом является присоединение молекулы малонил АПБ. к карбоксильному концу растущей цепи жирной кислоты.
Суммарное уравнение синтеза пальмитиновой кислоты;
СН3СОКоА+7СООНСН2СОSКоА+14НАДФН2
С15Н31СООН+7СО2+8НSКоА+14НАДФ+ 6Н2О
Ненасыщенные жирные кислоты в организме не синтезируются и поступают с пищей и их называют незаменимыми жирными кислотами. Пальмитолеиновая, олеиновая кислоты синтезируются из пальметиновой и стеариновая кислоты с участием оксигеназы и молекулярного кислорода и НАДФН2 .
Синтез триглицеридов
Для синтеза триглицеридов необходимы: Глицерин, жирные кислоты.
Глицерин образуется при распаде углеводов и липидов
14

Синтез фосфолипидов
Фосфолипиды синтезируются в печени, слизистой кишечника, почках, надпочечниках, нервной ткани.
Синтез фосфолипидов до образование фосфатидной кислоты идет также как и синтез триглицеридов.
Фосфотидная к-ка может идти либо на синтез триглециридов или на синтез фосфолипидов.
Зависит это от присутствия азотистых оснований (холин, серин, этаноламин) и ненасыщенных жирных кислот липотропные вещества метеомин, В12, они являются донорами метильних групп для образования холина.
Если много ненасыщенных жирных кислот и азотистых оснований синтезируются фосфолипиды, если их мало синтезируются триглицериды
15
Дипальмитоил фосфатидилхолин основной компонент сурфактанта легких. Сурфактант внеклеточный липидный слой с небольшим количеством гидро – фобных белков выстилающий поверхность альвеол и предотвращающий слипание стенок альвеол во время вдоха. Недостаточное формирование сурфактанта у недоношенных детей после рождении приводит к развитию респираторного дистресс – синдрома основной причины смерти у этой группы новорожденных.
Литература:
А. Основные: 1. Биохимия. Т.Т.Берѐзов, Б.Ф.Коровкин. 2010 2. Биохимия. Б.А.Строев. 1986
Б. Дополнительные:
1. Медицинская биохимия. С.М.Рапапорта. 1976 2.Биохимия. Р.Страер. 1985 3.Северин. 2003
16
Лекция № 4 Обмен холестерина
План:
1.Переваривание холестерина;
2.Синтез эндогенного холестерина;
3.Регуляция липидного обмена;
4.Нарушения липидного обмена
Пищевые источники –мясо, желток яйца, печень, мозг В организм человека ежедневно поступает 0,1-0,3г холестерина.
Холестерин в ЖКТ не подвергается расщеплению, холестериды расщепляютя в duodenum под действием холестеразы на холестерин и высшую жирную кислоту.
Холестерин в стенке тонкого кишечника всасывается с участием желчных кислот и в составе хиломикронов транспортируются кровью в печень.
80% экзогенного холестерина идет на синтез желчных кислот, гормонов надпочечников, половых гормонов. 20% экзогенного холестерина выводится из организма в виде копростерина.
Синтез эндогенного холестерина
Эндогенной холестерин синтезируется печени (более 50%), в тонком кишечнике (15-20%), остальной холестерин синтезируется в коже, коре надпочечников, половых железах. В сутки в организме синтезируется около 1г холестерина; с пищей поступает 300-500мг.
Синтез холестерина происходит в цитозоле клеток. Это один из самых длинных метаболических путей в организме человека. Сложный путь синтеза холестерина можно разделить на 3 этапа.
Первый этап – превращение ацетил КоА в мевалоновую кислоту. Второй этап – образование сквалена.
Третий этап – циклизация сквалена и образование холестерина.
Две молекулы ацетил КоА конденсируются ферментном тиолазой с образованием ацетоацетил КоА. Затем присоединяется еще присоединяется одна молекула ацетил КоА и образуется β – гидрокси β – метилглутарил КоА.
17
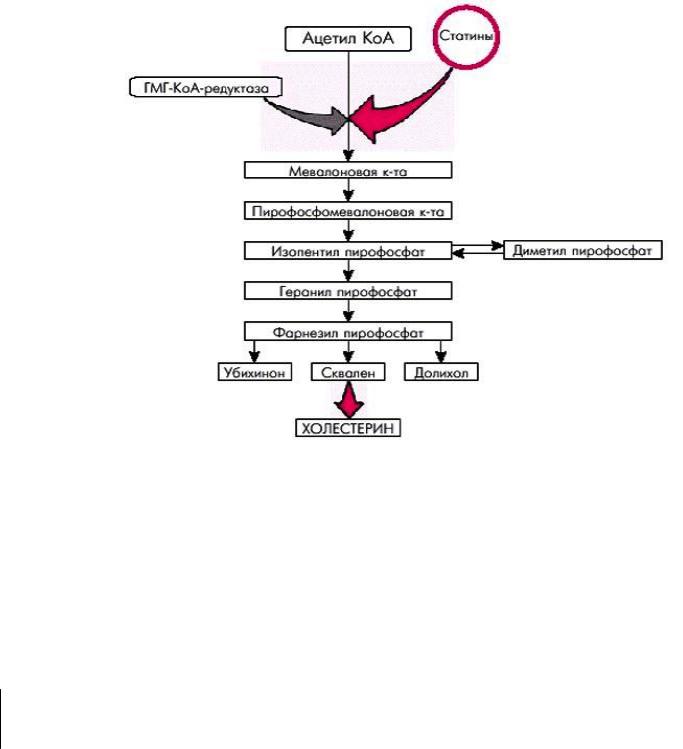
На втором этапе синтеза мевалоновая кислота превращается в пятиуглеродную изопреноидную структуру, содержащую пирофосфата. Продукт конденсации 2 изопреновых единец – геранилпирофосфата, затем фарнезилпирофосфата, состоящего из 15 углеродных атомов. Две молекулы фарнезилпирофосфат конденсируются с образованием сквалене, состоящего из за углеродных атомов.
На третьем этапе сквален превращается в ланостерин. Далее происходит 20 последовательных реакций, превращающих ланостерин в холестерин, содержащий 27 углеродных атомов.
Регуляция липидного обмена
Липидный обмен регулируется ЦНС и железами внутренний секреции. Кора мозга оказывает трофическое влияние на жировую ткань либо через симпатическую и парасимпатическую систему либо через эндокринные железы.
Известно, что длительный эмоциональный стресс, сопровождается вопросом катехоламин в кровяное русло и может вызвать похудание. Жировая ткань иннервируется симпатический и парасимпатической нервной системой и возбуждение сопровождается выделением адреналином и норадреналином, который увеличивает скорость липолиза в жировой ткани в результате усиливается мобилизация жирных кислот из жировой ткани.
18

Аденилат
адреналин |
и АМФ |
протеинкиназа |
триглицерид |
циклаза
липаза
 глицерин + жирные кислоты
глицерин + жирные кислоты
Глюкагон действует также как катехоламины. Гормон перед ней доли гипофиза, гормон роста усиливает синтез аденилатциклазы и триглицеридлипазы.
Гипофункция гипофиза приводит от отложению жира, развивается гипофизарное ожирение.
Гиперфункция приводит к гипофизарной кахексии.
Тироксин, глюкокортикоиды – активируют аденилатциклазу, вызывают липолиз.
Инсулин – активирует гликолиз и тормозит освобождение жирных кислот, превращает глюкозу в жиры.
Синтез холестерина зависит от активности редуктазы β-окси β- метилглутарил КоА (регуляторного фермента). Считают, что ингибитором этого фермента является экзогенный холестерин.
Метаболизм ацетил КоА
Углеводы |
липиды |
аминокислоты |
Синтез ацетоновых тел
|
|
|
Ацетил КоА |
|
|
|
|
|
|
|
|
|
|
|
НАДФН2 |
НАДФН2 |
|
ЦТК |
синтез |
|
синтез |
||
СО2 |
|
|
высших |
|
холестерина |
|
|
||||
Н2О |
|
|
жирных |
|
|
|
|
|
|||
АТФ |
кислот |
|
|
||
Синтез ацетоновых тел
Ацетоновые тела синтезируются из ацетил КоА
19

К кетоновым телам относятся: оксибутират, ацетоацетат и ацетон.
|
|
|
|
мозг |
|
|
|
|
|
|
|
|
-гидроксибутират |
||
|
|
|
|
|
|
|
|
|
|
||||||
|
|
|
|
сердце |
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
||||||
|
|
|
|
мышцы |
|
|
|
||||||||
|
|
|
|
|
|
||||||||||
ацетил КоА |
|
ацетоацетил КоА |
|
|
|
|
|
|
ацетоацетат |
||||||
|
|
|
|
|
|
||||||||||
|
|
|
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
|
|
|
|||||||
жирные к-ты |
|
|
жировая ткань |
|
|
|
глюкагон |
||||||||
|
|
|
|
|
|||||||||||
Нарушения липидного обмена могут наступать уже в процессе переваривания и всасывания липидов из-за недостаточного поступления панкреатической липазы в кишечник, из-за нарушения поступления в кишечник желчи.
К нарушениям липидного обмена относятся заболевания печени (жировая инфильтрация печени), при которой в печени откладывается большое количество триглицеридов. В норме в печени содержится 2-5 % Тг, при жировой инфильтрации содержания ТГ возрастает до 50-60%. Это может быть при сахарном диабета, углеводном голодании, когда ткани испытывают дефицит энергии. Происходит усиленный распад липидов, которые поступают в печень, но окисление нарушается из-за нарушения ферментов β- окисления.
20