

2 курс / Нормальная физиология / Руководство_к_практическим_занятиям_по_нормальной_физиологии_Правдивцев
.pdfКлетка А принимает поток входных импульсов, передавая их на нейрон В, который работает в паре с тормозным нейроном С. Что делает клетка В? Она отвечает на 1-й, 2-й, 3-й импульсы, поступающие к ней от нейрона А. Однако параллельно возбуждение идет к тормозному нейрону С. Нейрон С возбуждается, при этом тормозный медиатор в окончаниях аксона С тормозит активность нейрона В. Нейрон В перестает реагировать на импульсацию от нейрона А, следовательно, в структуре выходного потока клетки В появляется пауза. Однако, как только клетка В перестанет работать под влиянием нейрона С, последний сам перестает работать, т. к. к нему перестает поступать возбуждающая импульсация от нейрона В. Это немедленно восстанавливает возбудимость нейрона В, и он вновь начинает реагировать на импульсацию от нейрона А. Далее циклы повторяются, при этом выходной поток НЦ, определяемый активностью нейрона В, будет носит форму, в которой вспышки активности чередуются с тормозными паузами (рис. 9). В целом выходная импульсация НЦ будет редуцирована в сравнении с исходной.
Рис. 10. Схема нейронной сети НЦ с тормозным нейроном, включенным по схеме параллельного торможения.
Еще один пример с участием тормозного нейрона. Что будет на выходе НЦ, если тормозная клетка включается в нейронную сеть по схеме параллельного торможения?
Под влиянием начальной порции возбуждения выходной нейрон В генерирует несколько импульсов. Однако через коллатераль аксона А параллельно активируется тормозный нейрон С, который тормозит клетку В. Ее активность тормозится. В итоге – на выходе наблюдаем всего два импульса в сравнении с шестью ПД на входе (рис. 10).
5. Нервным центрам (НЦ) присуще явление последействия, под которым понимают повышенную активность нейронов НЦ на протяжении некоторого времени после прекращения поступления нервных импульсов на вход НЦ.
Как этот феномен объяснить с позиций нейрофизиологии? Линейная цепь нейронов А-В последействие не обеспечивает. Однако ситуация изменяется, когда между нейронами А и В встраивается замкнутая сеть вставочных нейронов 1, 2, 3 (рис. 11).
Рис. 11. Схема нейронной сети, обеспечивающей феномен последействия в НЦ за счет циркуляции (реверберации) нервных импульсов в цепи вставочных нейронов 1, 2, 3, замыкающихся на нейрон А. Цепь вставочных нейронов загружается импульсацией при поступлении к нейрону А входной (афферентной) импульсации (1). В дальнейшем (2) входная импульсация воспроизводится непрерывно в замкнутой цепи нейронов 1, 2, 3, далее нейрон А и выходной нейрон В.
От аксона А коллатераль поступает к нейрону 1, от нейрона 1 аксон поступает к нейрону 2, от нейрона 2 – к нейрону 3, а от нейрона 3 – к входному нейрону А. Формируется «ловушка возбуждения» – циклически замкнутый нейронный комплекс. Возбуждение в таком комплексе может длительно циркулировать, сохраняя рисунок входной импульсации, непрерывно поддерживая высокий уровень активности выходного нейрона В даже после прекращения поступления входной импульсации на нейрон А.
Учитывая, что «ловушки возбуждения» сохраняют рисунок входной импульсации (фактически запоминают параметры входной информации) в настоящее время их рассматривают в качестве одного из механизмов кратковременной памяти.
6. Нервные центры обладают способностью суммировать возбуждения, поступающие к ним по афферентным каналам. Суммация может быть двух видов: временная и пространственная.
Временная, или частотная суммация (рис. 12). К нейрону (А) НЦ в результате воздействия на входной нерв подходят друг за другом два нервных импульса. Каждый из импульсов вызывает генерацию ВПСП, недостаточную для выведения мембранного
21

потенциала нейрона до КУД (В). Однако если второй импульс приходит к нейрону на фоне развития ВПСП от первого, ВПСП от второго импульса, суммируясь с ВПСП от первого, смещает мембранный потенциал до КУД, нейрон НЦ при этом возбуждается. В рассмотренном примере суммация двух входных подпороговых воздействий к НЦ является производным фактора времени между входными воздействиями. При некотором оптимальном временном интервале два подпороговых воздействия могут суммироваться на нейронах НЦ.
Рис. 12. Временная (частотная) суммация входных воздействий на нейроне НЦ.
Пространственная суммация (рис. 13). К нейрону (А) по различным входным нервам поступают два нервных импульса. Каждый вызывает подпороговый ВПСП (В). Однако если нервные импульсы поступают одновременно – подпороговые ВПСП суммируются (С), при этом мембранный потенциал нейрона смещается до КУД, что приводит к возбуждению нейрона НЦ.
Рис. 13. Пространственная суммация входных воздействий на нейроне НЦ, когда два импульса подходят к нейрону одновременно по различных афферентным проводникам (феномен конвергенции).
7. Нервные центры (НЦ) обладают способностью перестраивать свою работу в зависимости от изменения активности сопряженных нервных центров, изменения качества афферентных воздействий на НЦ. Данное свойство НЦ определяется как пластичность.
Как доказать феномен пластичности НЦ? На рис. 12-1 представлена упрощенная схема моторных центров спинного мозга А и В, осуществляющих контроль за активностью периферических мышц конечностей А и В. Перережем эфферентные (моторные) нервы и соединим их по схеме – нерв А – с мышцами В, нерв В с – мышцами А (рис. 12-2). Спустя некоторое время функциональная целостность нервов восстанавливается, однако моторная активность подопытного животного после вмешательства грубо нарушается. Животное теряет способность нормально передвигаться. Тем не менее, после многочисленных попыток перемещения в среде обитания ситуация постепенно начинает улучшаться вплоть до полного восстановления в результате радикальной перестройки нейронной активности спинальных двигательных центров.
Рис. 12. Упрощенная схема моторных центров спинного мозга интактного животного (1) и животного после перекрестного сшивания эфферентных нервов (2).
22

Рис. 13. Кора головного мозга играет главную роль в обеспечении пластических реакций ЦНС после повреждений.
В рассмотренном примере пластические перестройки в работе спинальных нервных центров происходят преимущественно за счет пластических перестроек нейронной активности на уровне коры головного мозга. Именно нисходящие корковые возбуждения нового качества вносят решающий вклад в изменение работы спинальных нервных центров. Ведущую роль коры головного мозга в обеспечении пластических перестроек спинальных центров можно доказать в эксперименте. Перережьте нисходящие в спинной мозг корковые двигательные тракты у животного после восстановления двигательных функций – ситуация немедленно вернется к исходной.
8.Нервные центры (НЦ) обладают повышенной утомляемостью. Известно, что нервное волокно практически не утомляется (Н. Е. Введенский), напротив, НЦ обладают повышенной утомляемостью. Неслучайно в целом организме утомление раньше всего развивается в НЦ (И. М. Сеченов).
9.Нервные центры обладают повышенной чувствительностью к ядам, биологически активным веществам, лекарственным препаратам. Это связано с обилием синаптических контактов в структуре нейронных сетей НЦ, наличием множества химических механизмов передачи возбуждения через синапсы.
10.Нервные центры обладают повышенной чувствительностью к недостатку кислорода.
11.Нервные центры (НЦ) могут пребывать в трех функциональных состояниях, отражающих совокупный уровень активности всех нейронов НЦ. Исходное состояние НЦ – нормальная (обычная) активность нейронов НЦ; второе состояние – сниженная активность нейронов НЦ; третье состояние – повышенная активность нейронов НЦ. В последнем случае НЦ в составе ЦНС становится доминантным, или господствующим (А. А. Ухтомский).
Причины перехода НЦ из «обычного» состояния в доминантное состояние.
1. Гормональные причины, например, избыток или недостаток в циркулирующей крови тех или иных гормонов. 2. Патологические причины, например, хроническая болевая импульсация, формирующая доминантное состояние центров, ответственных за переработку болевой импульсации (ядра ретикулярной формации, гипоталамуса, таламуса, лимбической системы). 3. Метаболические причины, например, недостаток воды в организме, формирующий доминантное состояние гипоталамического центра жажды. 4. Информационные причины (применительно к человеку), связанные с восприятием словесных сигналов, письменных сообщений и проч., формирующих доминантное состояние корковых центров, имеющих первостепенное значение для организации целенаправленных поведенческих актов.
Доминантный НЦ обладает рядом специфических свойств. Среди них – стойкость, повышенная возбудимость нейронов.
Повышенная возбудимость доминантного НЦ предопределяет способность нейронов доминантного НЦ реагировать на возбуждения, на которые ранее они не реагировали (доминантный НЦ «притягивает к себе» иррадиирующие по ЦНС возбуждения).
На центры-конкуренты доминантный НЦ оказывает угнетающее влияние (например, человек в состоянии сильной жажды обычно отказывается от еды – доминантный центр жажды угнетает пищевой центр). Доминантный НЦ всегда активирует аппарат биологически отрицательных эмоций, поэтому доминантное состояние того или иного НЦ у человека всегда протекает на фоне неприятных, тягостных ощущений, переживаний. Последнее позволяет сделать вывод о том, что состояние доминанты в ЦНС формируется как реакция на возникновение жизненно-важных проблем, конкретных трудностей. Значение доминантного состояния НЦ заключается в том, что доминанта инициирует целенаправленный тип поведения, ориентированный на достижение полезных результатов, обеспечивающих устранение причин, вызвавших переход НЦ из обычного (нормального) состояния в состояние доминантное.
Координационная деятельность нервных центров
Функцию нервных центров в целом можно определить как координационную. Координационная деятельность НЦ – деятельность, направленная на обеспечение упорядоченности, согласованности в работе периферических эффекторов, активность которых поддерживает стабильность жизненно важных параметров внутренней среды организма в условиях постоянно изменяющейся среды обитания. Координационная деятельность НЦ реализуется в составе функциональной системы (рис. 14), под которой понимают центрально-периферическую организацию, обеспечивающую достижение полезных для организма конечных результатов. Таким образом, субстратом координационной деятельности НЦ является функциональная система.
В состав ФС (рис. 14) входят – нервный центр ( 1). Проводники, связывающие НЦ с периферическими эффекторами (канал прямой связи – 2). Совокупность периферических эффекторов (3), определяющих статус регулируемой константы (4). Рецептор регулируемой константы (5), активность которого отражает ее возможные отклонения от оптимального уровня. Канал обратной
23
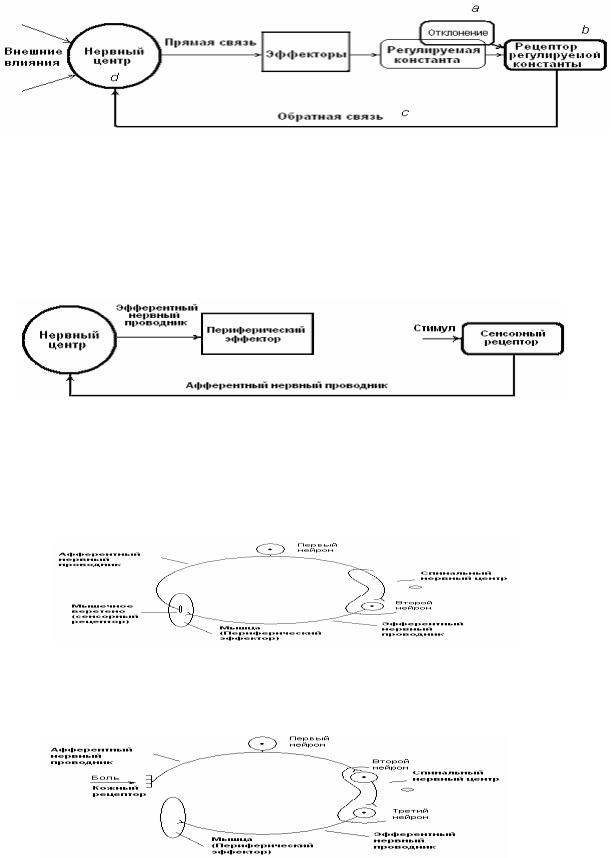
связи (6), по которому информация о статусе регулируемой константы поступает в НЦ, координирующий работу эффекторов,
обеспечивая стабильность регулируемой константы.
Рис. 14. Схема функциональной системы как субстрата координационной деятельности НЦ. a, b, c, d – последовательность событий в составе функциональной системы, направленных на предотвращение отклонения регулируемой константы от оптимального уровня (описание в тексте).
Вопрос. Какой элементарный механизм функциональной системы обеспечивает процесс вовлечения нервного центра (НЦ) в координационную деятельность?
Ответ. Механизм рефлекса, под которым понимают реакцию НЦ, проявляющуюся в изменении физиологической активности эффекторов функциональной системы, под влиянием афферентной импульсации. Путь, который проходит нервный импульс в процессе реализации рефлекса, получил название рефлекторной дуги. В состав рефлекторной дуги (рис. 15) входят: сенсорный рецептор, афферентные нервы, НЦ (нейроны НЦ), эфферентный нерв, периферический эффектор (эффекторный орган).
Рис. 15. Принципиальная схема дуги рефлекса как элементарного механизма функциональной системы, обеспечивающего в ответ на действие стимула (раздражителя) вовлечение нервного центра в координационную деятельность по отношению к периферическому эффектору или эффекторам.
Примеры простейших рефлексов (рефлекторных актов).
Миотатический рефлекс (коленный рефлекс) – рефлекс в ответ на растяжение мышцы (мышечных веретен), проявляющийся в том, что нейроны спинального НЦ вызывают сокращение этой же мышцы. Схема рефлекторной дуги, включающая всего два нейрона
(моносинаптический рефлекс), представлена на рис. 16.
Рис. 16. Схема двухнейронной рефлекторной дуги миотатического рефлекса.
Сгибательный рефлекс – рефлекс в ответ на раздражение кожных рецепторов, проявляющийся в том, что нейроны спинального нервного центра вызывают сокращение сгибательных мышц конечности. Схема рефлекторной дуги, включающая три нейрона
(полисинаптический рефлекс), представлена на рис. 17.
Рис. 17. Схема трехнейронной рефлекторной дуги сгибательного рефлекса.
Простая классификация рефлексов (рефлекторных актов).
24
1.Рефлексы классифицируются по критерию локализации сенсорных рецепторов, возбуждение которых вызывает тот или иной рефлекс. Условно всю совокупность рецепторов делят на экстеро- , интеро- и проприорецепторы. Следовательно, рефлексы можно подразделить на а) экстероцептивные рефлексы; б) интероцептивные рефлексы; в) проприоцептивные рефлексы.
2.Рефлексы классифицируются по критерию локализация нервных центров, обеспечивающих соответствующие рефлексы. Отсюда
– а) спинальные рефлексы, реализуемые спинным мозгом; б) бульбарные рефлексы, реализуемые продолговатым мозгом; в) мезенцефальные рефлексы, реализуемые средним мозгом; г) диэнцефальные рефлексы, реализуемые промежуточным мозгом; д) кортикальные рефлексы, реализуемые корой головного мозга.
3.Рефлексы классифицируются по числу синаптических переключений. Соответственно различают – а) моносинаптические рефлексы (один синаптический контакт в составе двухнейронной рефлекторной дуги; б) полисинаптические рефлексы с множеством синаптических переключений в нервном центре.
4.Рефлексы классифицируются по характеру рефлекторного эфферентного процесса. По данному критерию различают: а) двигательные рефлексы, если в конце рефлекторного акта сокращается мышца; б) секреторные, если в конце – секреция железы; в)
сердечные; г) желудочные, д) сосудистые и т.д.
Указанные критерии используются для идентификации рефлекторных актов. Например, |
рефлекс, вызывающий сгибание |
конечности при раздражении сенсорных кожных рецепторов – это экстероцептивный, |
спинальный, полисинаптический |
двигательный рефлекс. |
|
Идея рефлекса была сформулирована Рене Декартом – французским естествоиспытателем и математиком (XVII век). Термин «рефлекс» в научную литературу ввел Ян Прохазка - чешским ученым (XIX век). Особое развитие рефлекторная теория получила в России благодаря работам И. М. Сеченова и И.П. Павлова. И. П. Павлов впервые сформулировал принципы рефлекторной теории:
1. Принцип детерминизма (причинности), 2.Принцип структурности. 3.Принцип единства процессов анализа и синтеза в рамках рефлекторных актов.
Частные принципы координационной деятельности нервных центров (НЦ).
1.Принцип прямой связи (рис. 14) НЦ с эффекторами. Прямая связь предоставляет НЦ реальную возможность координировать работу эффекторов функциональной системы.
2.Принцип обратной связи. По каналам обратной связи НЦ получает информацию о достаточности или недостаточности координирующих влияний на периферические эффекторы в рамках регуляции по отклонению, при которой первичным пусковым фактором, активирующим новую координацию, выступает отклонение регулируемой константы от заданного уровня. Например, в силу каких-то причин у нормального человека температура внутренней среды обнаружила тенденцию к увеличению. В этом случае немедленно активируются системные рецепторы константы. По каналам обратной связи информация поступает в НЦ, который,
изменяя качество координации периферических эффекторов, предотвращает сдвиг температурной константы в сторону увеличения (рис. 14).
3.Принцип конвергенции входных возбуждений на нейронах НЦ. Под конвергенцией понимают схождение разнородных возбуждений к одним и тем же нейронам. В результате конвергенции происходит интеграция (объединение) множественных потоков входных возбуждений в один выходной, как это показано на рисунках 2, 13.
4.Принцип реципрокности, или исключительности в работе нейронных сетей нервных центров (НЦ). Различные НЦ в составе ЦНС, как правило, состоят из двух «антагонистических» подцентров. Например, дыхательный центр включает подцентры вдоха и выдоха; терморегуляторный центр – подцентры теплопродукции и теплоотдачи; сосудодвигательный центр – подцентры прессорный и депрессорный; гипоталамический пищевой центр – подцентры голода и насыщения. Возникает вопрос – на основе какого принципа соотносится деятельность подцентров-антагонистов, реализующих принципиально различные эффекты на периферии? Ответ – на основе принципа реципрокности, или исключительности (рис. 15).
На рис. 18 представлена схема, иллюстрирующая принцип реципрокности применительно к активности спинального моторного центра, обеспечивающего сгибание и разгибание конечности, например при ходьбе. Отметим, что спинальный нервный центр состоит из двух подцентров. С одной стороны – это подцентр, контролирующий активность сгибательных мышц в составе входного нейрона А и моторного нейрона B, с другой – подцентр, контролирующий активность разгибательных мышц в составе входного нейрона С и моторного нейрона D. Как будет работать центр в ситуации, когда конечность должна согнуться? Для обеспечения результативного сгибания необходимо, чтобы в первую очередь сокращались сгибательные мышцы, вместе с тем, в этот же самый момент необходимо, чтобы разгибательные мышцы пребывали бы в состоянии расслабления.
Исключительность в работе мышц-антагонистов (одни сгибаются, другие расслабляются) обеспечивается реципрокными взаимоотношениями в работе сгибательного и разгибательного подцентров. При этом, если афферентное возбуждение (1) первично поступает к нейронам сгибательного подцентра (нейроны А и В ), обеспечивающих сгибание, по коллатерали нейрона А возбуждается тормозный нейрон Y в составе разгибательного подцентра, при этом выходной – моторный нейрон D переходит в состояние торможения. Иначе, в ответ на возбуждение 1 нейрон В – возбуждается, сгибатели сокращаются, тогда как моторный нейрон D – тормозится, разгибатели расслабляются. Все вместе взятое обеспечивает адекватный процесс сгибания конечности. Если ситуация меняется, и в спинальный нервный центр первично поступает возбуждение 2, возбуждаться будет нейрон D, тогда как нейрон В будет тормозиться, конечность в этой ситуации будет разгибаться.
25

Рис. 18. Схема, иллюстрирующая принцип реципрокности в работе моторного спинального центра, обеспечивающего процессы сгибания и разгибания конечности.
ТЕМА СЕРДЦЕ. ФАЗЫ СЕРДЕЧНОГО ЦИКЛА. ОСНОВНЫЕ ФИЗИОЛОГИЧЕСКИЕ СВОЙСТВА СЕРДЕЧНОЙ МЫШЦЫ. (стр. 271-279)
Контрольные вопросы. Строение сердца, значение камер, клапанов. Круги кровообращения. В каком положении находятся клапаны сердца в момент, предшествующий началу кардиоцикла? Чему равна продолжительность систолы желудочков? Из каких периодов она состоит? Из каких фаз складывается период напряжения, укажите продолжительность каждой фазы? Из каких фаз складывается период изгнания крови, укажите продолжительность каждой фазы? Почему
при |
сокращении |
желудочков |
створки |
атриовентрикулярных клапанов не |
|
выворачиваются |
в |
предсердия? Чему равна продолжительность диастолы желудочков? Что |
|||
такое протодиастолический период, чему он равен? |
Чему равна продолжительность систолы |
||||
предсердий? Чему равна продолжительность диастолы предсердий? Чему равна длительность кардиоцикла, если частота сокращений сердца составляет 70/мин? Чему равна частота сердечных сокращений, если длительность кардиоцикла составляет 1 сек? Нарисуйте
механокардиограмму, |
обозначьте |
интервал, |
соответствующий кардиоциклу? |
Нарисуйте |
||
механокардиограмму, |
обозначьте интервал, |
соответствующий систоле и диастоле желудочков? |
||||
Нарисуйте механокардиограмму, |
обозначьте |
интервал, соответствующий систоле и диастоле |
||||
предсердий? Нарисуйте механокардиограмму, обозначьте момент закрытия и |
открытия |
|||||
атриовентрикулярных клапанов. Нарисуйте |
механокардиограмму, |
обозначьте |
момент, |
|||
соответствующий открытию и закрытию полулунных клапанов полулунных клапанов. Какие
клапаны закрыты большую часть сердечного цикла? Какие клапаны |
сердца открыты |
большую |
|||
часть сердечного |
цикла? |
|
|
|
|
РАБОТА 1. Регистрация механокардиограммы. Фазы |
сердечной |
деятельности. |
Смотри |
||
«Руководство» - работа 5.1. С. 179. |
|
|
|
|
|
Указания к оформлению протокола - |
|
|
|
|
|
1. Зарисуйте |
механокардиограмму, |
стрелками |
обозначьте |
основные фазы сердечной |
|
деятельности. |
|
|
|
|
|
2.Рассчитайте время основных фаз сердечного цикла - систолы и диастолы предсердий, систолы и диастолы желудочка, а также время кардиоцикла.
3.Определите частоту сокращения сердца лягушки исходя из продолжительности кардиоцикла.
4.Что отражает механокардиограмма?
--------------------------------------------------------------
Протокол
26
РАБОТА 2. Расчет продолжительности сердечного цикла по частоте пульса в покое и после физической нагрузки у человека.
Указания по выполнению работы и оформлению протокола
1.Подсчитайте по пульсу частоту сердечных сокращений за 1мин в покое.
2.Рассчитайте длительность сердечного цикла в покое.
3.Сделайте 20 приседаний с выбросом рук вперед за 30 с.
4.Рассчитайте продолжительность сердечного цикла после нагрузки.
5.Сравните результаты, сделайте выводы об изменении длительности кардиоцикла после физической нагрузки.
--------------------------------------------------------------
Протокол
Тема зачтена
ТЕМА – СВОЙСТВА СЕРДЕЧНОЙ МЫШЦЫ. ВОЗНИКНОВЕНИЕ |
И |
ПРОВЕДЕНИЕ |
|||
ВОЗБУЖДЕНИЯ В СЕРДЦЕ. АВТОМАТИЯ СЕРДЦА. (стр. 279-282) |
|
|
|
||
Контрольные вопросы. Какими свойствами обладает сердечная мышца? |
Что такое нексус? |
||||
Нарисуйте потенциал действия волокон миокарда, обозначьте |
его фазы? Чем отличается |
||||
проведение |
возбуждения по волокнам сердечной мышцы от |
проведения |
возбуждения |
по |
|
волокнам |
скелетной мышцы? Чем объясняется сокращение |
миокарда |
на |
пороговые |
и |
сверхпороговые раздражители по закону "все или ничего"? Какие ионные токи обеспечивают потенциал действия миокарда? Как можно доказать способность сердца к автоматии? Охарактеризуйте процесс распространения возбуждения от синусного узла до миокарда желудочков. Каковы функции проводящей системы сердца? Какова функциональная роль атриовентрикулярной задержки? Какая электрофизиологическая особенность свойственна клеткам
проводящей системы? Чем отличается деятельность сердца, работающего в синусном |
ритме от |
|||
деятельности сердца, |
работающего |
в |
атриовентрикулярном ритме? Как доказать, что в |
|
норме водителем ритма сердца является |
синусный узел? Нарисуйте |
изменения |
||
механокардиограммы |
после наложения |
|
лигатуры, изолирующей синусный узел? Что такое |
|
преавтоматическая пауза? Как можно |
уменьшить продолжительность преавтоматической |
|||
|
|
|
27 |
|
паузы? Как доказать, что типичным миокардиальным клеткам автоматия не свойственна? Что такое экстрасистола?
РАБОТА 1. Роль синусного узла в автоматии сердца (опыт Гаскелла ) Откройте сердце у лягушки, соберите установку для записи
механокардиограммы (МКГ). Запишите МКГ на медленно вращающемся барабане кимографа. Подсчитайте частоту сердечных сокращений. Не останавливая кимограф, приложите к области синусного узла дно пробирки, заполненной льдом. Этот момент отметьте на МКГ стрелкой. Когда на МКГ появится эффект, пробирку уберите, вновь подсчитайте частоту сердечных сокращений. После возвращения МКГ к норме проверьте эффект нагревания синусного узла. Опыт проводится так же, но в пробирку наливается вода порядка 30-36 град. Аналогичным образом проверьте действие охлаждения и нагревания миокарда желудочка.
Указания к оформлению протокола -
1.Зарисуйте изменения МКГ при охлаждении и нагревании области синусного узла.
2.Зарисуйте изменения МКГ при охлаждении и нагревании миокарда желудочков.
3.Объясните полученные результаты работы.
--------------------------------------------------------------
Протокол
РАБОТА 2 .Анализ проводящей системы сердца (опыт Станниуса). Смотри «Руководство» -
работа 5.5. С. 183.
Указания к оформлению протокола -
1.Зарисуйте механокардиограмму после наложения 1 лигатуры по Станниусу. Объясните, почему не сокращаются предсердия и желудочек.
2.Зарисуйте механокардиограмму после наложения 2 лигатуры по Станниусу. Объясните
причину возобновления деятельности сердца. Чем отличается работа сердца после наложения 2 лигатуры по Станниусу от исходной работы.
3.Что доказывает опыт наложения 3 лигатуры по Станниусу.
4.Что понимают под градиентом автоматии?
--------------------------------------------------------------
Протокол
РАБОТА 3. Особенности возбудимости сердца. Экстрасистола. Смотри «Руководство» - работа
5.6. С. 185.
28
Одной из важнейших физиологических особенностей |
сердечной мышцы является длительный |
|||||||||||||
рефрактерный период, который состоит из |
фазы |
абсолютной |
и |
|
фазы |
относительной |
||||||||
рефрактерности. Абсолютная рефрактерная фаза по времени почти |
|
совпадает с систолой, |
||||||||||||
относительная рефрактерная фаза - с диастолой. Если во время |
систолы |
желудочка |
раздражать |
|||||||||||
его |
одиночными импульсами сверхпорогового |
электрического тока, |
|
можно убедиться, что |
||||||||||
раздражение остается без ответа. |
Раздражение, |
нанесенное на |
желудочек |
во время |
диастолы, |
|||||||||
вызывает внеочередное сокращение - экстрасистолу, амплитуда которой тем больше, |
чем |
ближе |
||||||||||||
к |
концу диастолы наносится |
второе раэдражение. |
За желудочковой |
экстрасистолой |
всегда |
|||||||||
следует более |
продолжительная |
чем обычно |
пауза, |
называемая компенсаторной паузой |
||||||||||
Указания к оформлению протокола - |
|
|
|
|
|
|
|
|
|
|
||||
1. Нарисуйте |
|
механокардиограмму |
при |
нанесении сверхпорогового |
электрического |
|||||||||
раздражения |
на |
миокард желудочков во время систолы. |
Объясните |
причину |
отсутствия |
|||||||||
изменений хода механокардиограммы. |
|
|
|
|
|
|
|
|
|
|
||||
2. Нарисуйте механокардиограмму при нанесении сверхпорогового |
|
электрического |
||||||||||||
раздражения |
на |
миокард желудочков |
во |
время |
диастолы. |
Отметьте |
на |
рисунке |
||||||
экстрасистолу |
и |
компенсаторную паузу. |
|
|
|
|
|
|
|
|
|
|
||
3.Объясните причину возникновения компенсаторной паузы после желудочковой экстрасистолы.
4.Нарисуйте график изменения возбудимости миокарда во время его сокращения и расслабления.
--------------------------------------------------------------
Протокол
Биофизические основы электрокардиографии
Особенности распространения возбуждения по сердцу отображаются в электрокардиограмме (ЭКГ), которая имеет характерную форму (рис. 4). Элементарной моделью генератора ЭКГ является электрический диполь. При распространении возбуждения по миокарду формируется множество диполей, которые закономерно изменяют свои количественные характеристики и направление. В каждый момент времени возникают новые диполи, исчезают прежние. В результате на поверхности сердечной мышцы создается сложная мозаика распределения электрических потенциалов. Результирующий дипольный момент миокарда, равный векторной сумме отдельных диполей, получил название интегрального электрического вектора сердца
(ИЭВС). Ритмичный характер автоматизма водителя ритма, а также передача возбуждения посредством электрических синапсов обусловливают синфазность возбудительного процесса в миокардиальных волокнах. Поэтому ИЭВС имеет сравнительно большую амплитуду прежде всего при деполяризации желудочков, чем создается высокий уровень биопотенциалов, отражающих сердечную деятельность даже на поверхности тела. Ежемоментно амплитуда и направление ИЭВС различны. Измеряя их, врач получает сведения о движении волны возбуждения по сердцу, что позволяет ему оценить свойства миокарда и в случае нарушений сердечной деятельности понять их природу.
29

Рис. 4. Записи ЭКГ в трех стандартных отведениях и схема, показывающая связь между распространением возбуждения в сердце и возникновением некоторых зубцов электрокардиограммы (по Барду).
При одновременной подаче электрических потенциалов, отражающих распространение возбуждения по миокарду, на пластины “Х” и “У” электроннолучевого осциллографа на его экране возникают фигуры Лиссажу, образующиеся в результате сложения взаимно перпендикулярных колебаний регистрируемых потенциалов (рис. 5).
Рис. 5. Соотношение векторэлектрокардиограммы (А) и электрокардиограммы (Б).
В данном случае фигуры Лиссажу представляют собой траектории движения ИЭВС, описываемые его концом на плоскости, перпендикулярной направлению распространения возбуждения по миокарду. Такое исследование электрической активности сердца называется векторэлектрокардиоскопией (ВЭКС). На векторэлектрокардиограмме выделяют обычно три эллипсоподобные фигуры. Самая мелкая из них отображает деполяризацию предсердий, самая крупная - деполяризацию желудочкой, средняя - их реполяризацию.
Амплитуду вектора оценивают посредством измерения его проекций на координатные оси. Любое
из отведений ЭКГ есть не что иное как проекция интегрального электрического вектора сердца на соответствующую координатную ось.
30