

2 курс / Нормальная физиология / Физиология_дыхания_Частоедова_И_А_,_Еликов_А_В_
.pdf71
5.7. Ингибиторы тканевого дыхания
Ингибиторы тканевого дыхания блокируют один из трех пунктов сопряжения дыхания и фосфорилирования. Все ингибиторы можно разделить на три группы, действие которых представлено на схеме 11.
I группа ингибирует дыхательную цепь на уровне 1-го пункта сопряжения дыхания и фосфорилирования. К данным веществам относятся, например,
ротенон («рыбий яд»), амобарбитал (препарат барбитурового ряда). При этом прерывается поступление водорода от субстратов окисляющихся через НАД-
зависимые дегидрогеназы. Выключается также первое из трех звеньев фосфорилирования. Данная группа ингибиторов не влияет на использование субстратов, окисляющихся через ФАД-зависимые дегидрогеназы.
|
Субстрат |
Ротенон |
ФАДН2 |
|
Антимицин А |
|
|
|
|
|
|||
|
|
|
|
|||
|
|
|
Амобарбитал |
Карбо- – |
|
БАЛ |
|
|
|
_ |
ксин |
|
|
|
|
|
|
|
|
|
|
НАДН•Н+ |
|
НАДН-дегидро- |
QH2 |
QH2-дегидрогеназа |
|
|
|
|
||||
|
|
|
||||
|
|
|
геназа ФМН |
|
в |
с |
H2S, CO, CN–
 –
–
 цитохром С
цитохром С  цитохромоксидаза
цитохромоксидаза  ½О2 а
½О2 а  а3
а3
Схема 11. Ингибирование ферментов цепи переноса электронов
II группа ингибиторов, к которым относится антимицин А (антибиотик,
выделенный из стрептомицетов) блокирует дыхательную цепь на уровне 2-го сопряжения дыхания и фосфорилирования и выключает участок цепи до блока.
|
|
|
|
|
|
|
|
72 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
O |
|
|
|
|
|
|
|
|
O |
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
C |
|
|
A |
AH2 |
|
|
|
C |
|
|
|
|||
|
|
|
|
|
|
|
|
|
|||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
HO |
|
|
|
|
|
|
O |
|
C |
||||||||
|
|
C |
|
|
|
|
|
|
|
|
O |
||||||
|
|
|
|
|
|
O |
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
HO |
|
|
|
|
|
|
|
|
O |
|
C |
||||||
|
|
C |
DH2 |
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
|
D |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
HC |
|
|
||||||
|
|
|
|
|
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
HC |
|
|
|
|
|
|
|||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
HO |
|
|
|
CH |
||||
HO |
|
|
CH |
|
|
|
|
||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
CH2OH |
|||||
|
|
|
CH2OH |
|
|
|
|
||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
восстановленная форма |
|
окисленная форма |
|||||||||||||||
аскорбиновой кислоты |
(дегидроаскорбиновая кислота) |
|
Схема 12. Функционирования окислительно-восстановительной пары с участием
аскорбиновой и дегидроаскорбиновой кислот
Дыхание возможно только в присутствии аскорбиновой кислоты, которая,
превращаясь в дегидроаскорбиновую кислоту по уравнению, представленному ниже, передает водород непосредственно на цитохром с. При этом образуется только одна молекула АТФ на уровне цитохромоксидазы.
III группа ингибиторов тканевого дыхания, к которым относятся цианиды
(например, NaCN, KCN), сероводород (H2S), оксид углерода (II), (СО, угарный газ) необратимо блокируют цитохромоксидазу, т.е. 3-й пункт сопряжения дыхания и фосфорилирования.
Это приводит к полному прекращению тканевого дыхания и быстрой гибели клетки от дефицита энергии. Поэтому ингибиторы цитохромоксидазы являются сильнейшими ядами. Оксид углерода (II), кроме того, нарушает транспорт кислорода кровью путем образования с гемоглобином устойчивого соединения
- карбоксигемоглобин.
Контрольные вопросы
1.На какие группы по механизму действия классифицируют ингибиторы ?
2.Какие ингибиторы относятся к I группе и как они действуют на цепь переноса электронов ?
73
3.Какие ингибиторы относятся ко II группе и как они действуют на цепь переноса электронов ?
4.Какие ингибиторы относятся к III группе и как они действуют на цепь переноса электронов ?
5.8. Тестовые задания и ситуационная задача
Выберите один правильный ответ.
26.ЦИАНИДЫ, УГАРНЫЙ ГАЗ, СЕРОВОДОРОД ЯВЛЯЮТСЯ ИНГИБИТОРАМИ ЦИТОХРОМОКСИДАЗЫ
1)аллостерического
2)конкурентного
3)необратимого действия
27.ДОНОРОМ ЭЛЕКТРОНОВ ДЛЯ УКОРОЧЕННОЙ ДЫХАТЕЛЬНОЙ ЦЕПИ ЯВЛЯЕТСЯ
1)пируват
2)малат
3)цитрат
4)сукцинат
5)б-кетоглутатат
28.ПРОСТЕТИЧЕСКОЙ ГРУППОЙ ПЕРВИЧНЫХ АКЦЕПТОРОВ ВОДОРОДА ФЛАВИНОВЫХ ДЕГИДРОГЕНАЗ ЯВЛЯЕТСЯ
1)НАДФ+
2)ФАД
3)ФМН
29.В СОСТАВ ПРОСТЕТИЧЕСКОЙ ГРУППЫ НАДН – ДЕГИДРОГЕНАЗНОГО КОМПЛЕКСА ВХОДИТ
1)ФМН
2)ФАД
3)хинон
30.В СОСТАВ ПРОСТЕТИЧЕСКИХ ГРУПП ФЛАВИНОВЫХ ДЕГИДРОГЕНАЗ ВХОДИТ ВИТАМИН
1)В1
2)В2
3)В5
4)В3
5)В6
74
31.КОЛИЧЕСТВО ЭНЕРГИИ, ВЫДЕЛЯЮЩЕЙСЯ ПРИ ПЕРЕНОСЕ
ЭЛЕКТРОНОВ ОТ ФАДН2 К МОЛЕКУЛЯРНОМУ КИСЛОРОДУ, ОБЕСПЕЧИВАЕТ СИНТЕЗ АТФ
1)3
2)2
3)1
32.ПРИ БЛОКИРОВАНИИ АМОБАРБИТАЛОМ ПЕРВОГО ПУНКТА ЦЕПИ ПЕРЕНОСА ЭЛЕКТРОНОВ СИНТЕЗ АТФ В МИТОХОНДРИЯХ ВОЗМОЖЕН ПРИ ИСПОЛЬЗОВАНИИ В КАЧЕСТВЕ СУБСТРАТА
1)малата
2)сукцината
3)б-кетоглутарата
4)глутамата
33.ДЫХАТЕЛЬНЫМ КОНТРОЛЕМ НАЗЫВАЕТСЯ РЕГУЛЯЦИЯ СКОРОСТИ ДЫХАНИЯ
1)цитохромоксидазой
2)НАДН-дегидрогеназой
3)концентрацией АДФ
Ситуационная задача 4
В суспензию митохондрий добавили 0,6 ммоль/л малата и 0,3 ммоль/л АДФ.
Поглощение кислорода, начавшееся сразу после добавления малата и АДФ,
через некоторое время прекратилось. Почему? Затем к пробе добавили раствор фермента гексокиназы и раствор глюкозы. Поглощение кислорода вновь началось. Почему?
Ситуационная задача 5
Сколько молекул АТФ образуется при окислении сукцината? Как будет влиять амобарбитал на окисление этого соединения?
6. Регуляция дыхания
Дыхание - одна из важнейших функций организма. Прекращение дыхания влечет за собой неминуемую гибель человека уже через 3-5 мин. Запасы кислорода в организме очень незначительны, поэтому необходимо его

75
постоянное поступление через систему внешнего дыхания. Указанное обстоятельство объясняет формирование в процессе эволюции такого механизма регуляции, который должен обеспечивать высокую надежность выполнения дыхательных движений. В основе деятельности системы регуляции дыхания лежит поддержание константного уровня таких показателей организма, как Рсо2, Ро2, рН. Их содержание остается относительно постоянным , несмотря на различные потребности в нем, которые во время интенсивной мышечной работы могут увеличиваться в 20 раз.
Регуляция дыхания направлена на выполнение двух задач: во-первых,
автоматическая генерация частоты и силы сокращения дыхательных мышц, во-
вторых, подстройка ритма и глубины дыхательных движений к реальным потребностям организма (в первую очередь, к изменениям метаболических параметров в виде рO2, рCO2 и pH артериальной крови и рCO2 и рH
межклеточной жидкости мозга).
Рис. 20. Нервный контроль вентиляции
76
Дыхание регулируется рефлекторным путем, включающим три составных элемента (рис. 20):
-рецепторы, воспринимающие информацию, и афферентные пути, передающие ее нервным центрам;
-нервные центры;
-эффекторы – пути передачи команд от центров и сами регулируемые объекты.
На рисунке 20 представлены образующие систему регуляции дыхания
блоки — контролирующие (нервные центры), исполнительные (дыхательные мышцы), рецепторные (хемо- и механорецепторы) и связи между этими блоками. Таким образом, вся система регуляции дыхания состоит из нескольких взаимосвязанных регуляторных контуров.
Входящие сигналы. Генератор ритма получает нисходящую из коры головного мозга импульсацию, а также нервные сигналы от нервных клеток интегратора сенсорной информации и непосредственно от центральных хеморецепторов.
Выходящие сигналы. Нервная импульсация от генератора ритма направляется к иннервирующим дыхательные мышцы двигательным нервным клеткам соответствующих ядер черепных нервов (VII, IX–XII) и к мотонейронам передних рогов спинного мозга (их аксоны в составе спинномозговых нервов направляются к дыхательным мышцам).
Интегратор сенсорной информации получает чувствительную информацию от разнообразных хемо- и механорецепторов, расположенных в органах дыхания и дыхательных мышцах, по ходу магистральных кровеносных сосудов
(периферические хеморецепторы), а также в продолговатом мозге (центральные хеморецепторы). Помимо этих прямых сигналов, интегратор получает множество информации, опосредованной различными структурами мозга (в
том числе и от высших отделов ЦНС). Импульсация от нервных клеток

77
интегратора, направляясь к нейронам генератора ритма, модулирует характер
разрядов от них.
Контрольные вопросы
1.Что лежит в основе деятельности системы регуляции дыхания ?
2.Какие составные элементы образуют систему регуляции дыхания ?
3.Какова роль интеграции сенсорной информации в системе регуляции дыхания ?
6.1.Дыхательный центр
Дыхательный центр - совокупность нейронных ансамблей разных этажей
центральной нервной системы, обеспечивающих управление внешним
дыханием (рис. 21).
КОРКОВЫЙ ОТДЕЛ ДЫХАТЕЛЬНОГО ЦЕНТРА
ЛИМБИЧЕСКИЙ ОТДЕЛ ДЫХАТЕЛЬНОГО ЦЕНТРА
ГИПОТАЛАМИЧЕСКИЙ ОТДЕЛ ДЫХАТЕЛЬНОГО ЦЕНТРА
ПНЕВМОТАКСИЧЕСКИЙ ЦЕНТР ВАРОЛИЕВА МОСТА
ДЫХАТЕЛЬНЫЙ ЦЕНТР ПРОДОЛГОВАТОГО МОЗГА
СПИНАЛЬНЫЕ МОТОНЕЙРОНЫ МЕЖРЕБЕРНЫХ МЫШЦ
Рис. 21. Уровни организации дыхательного центра
78
Главная часть дыхательного центра находится в продолговатом мозге. Это было установлено в опытах с перерезкой ствола мозга, проведенных французским физиологом Легуллуа в 1812 г. Миславский Н.А. (отечественный ученый) установил наличие двух структур, ответственных за вдох и выдох,
которые расположены в продолговатом мозге.
Автоматический дыхательный центр - совокупность нейронов специфических (дыхательных) ядер продолговатого мозга, способных генерировать дыхательный ритм (генераторы ритма).
Основная часть нейронов дыхательного центра продолговатого мозга относится к ретикулярной формации, эти нейроны обладают свойством спонтанной активности.
Дыхательный центр выполняет две основные функции в системе дыхания:
1)моторную, или двигательную, которая проявляется в виде сокращения дыхательных мышц
2)гомеостатическую, связанную с изменением характера дыхания при сдвигах содержания О2 и СО2 во внутренней среде организма.
Двигательная функция дыхательного центра заключается в генерации дыхательного ритма и его паттерна (длительность вдоха и выдоха). Моторная функция дыхательного центра адаптирует дыхание к метаболическим потребностям организма, приспосабливает дыхание в поведенческих реакциях
(поза, бег и др.), а также осуществляет интеграцию дыхания с другими функциями ЦНС.
Гомеостатическая функция дыхательного центра поддерживает нормальные величины дыхательных газов (O2, CO2) и рН в крови и внеклеточной жидкости мозга, регулирует дыхание при изменении температуры тела, адаптирует дыхательную функцию к условиям измененной газовой среды, например при пониженном и повышенном барометрическом давлении.
79
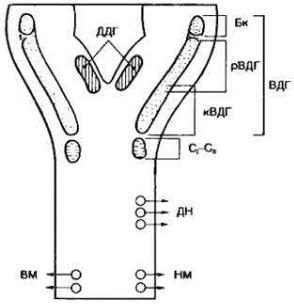
Нейроны дыхательного центра локализованы в дорсомедиальной и вентролатеральной областях продолговатого мозга и образуют так называемые дорсальную и вентральную дыхательную группу (рис. 22).
Дыхательные нейроны, активность которых вызывает инспирацию или экспирацию, называются соответственно инспираторными и экспираторными нейронами. Инспираторные и экспираторные нейроны иннервируют дыхательные мышцы.
Рис. 22. Проекция местоположения дыхательного центра на дорсальную поверхность продолговатого мозга. ДДГ и ВДГ — соответственно дорсальная и вентральная
дыхательные группы; Бк — комплекс Бетцингера; рВДГ и кВДГ — ростральная и каудальная часть ВДГ; СI—СII — сегменты спинного мозга; ДН, НМ и ВМ — соответственно диафрагмальный нерв и нервы наружных и внутренних межреберных мышц.
Дорсальная дыхательная группа (ДДГ) включает в себя симметричные области продолговатого мозга, расположенные вентролатеральнее ядра одиночного пучка (рис. 22).
Большинство нейронов этой группы (около 90 %) являются инспираторными. Нейроны ДДГ получают афферентные сигналы от легочных рецепторов растяжения по волокнам блуждающего нерва, нейроны которого
80
имеют обширные синаптические связи с другими отделами дыхательного центра и с различными отделами ЦНС. Только часть инспираторных нейронов ДДГ связана аксонами с дыхательными мотонейронами спинного мозга,
преимущественно с контралатеральной стороны. Другая часть нейронов взаимодействует с нейронами вентральной группы. Базовый ритм дыхания генерируется в основном дорсальной группой дыхательных нейронов. Даже после перерезки всех входящих в продолговатый мозг периферических нервов и ствола мозга выше и ниже продолговатого мозга эта группа нейронов продолжает генерировать потенциалы действия.
Вентральная дыхательная группа (ВДГ) расположена латеральнее обоюдного ядра продолговатого мозга, или ядра блуждающего нерва. ВДГ подразделяется на ростральную и каудальную части относительно уровня задвижки (obex) продолговатого мозга (рис. 22). Нейроны вентральной группы подразделяются на две части:
1)Ростральная часть ВДГ состоит из инспираторных нейронов, часть которых взаимодействует с другими дыхательными нейронами продолговатого мозга, другая часть нейронов направляют свои аксоны к дыхательным мотонейронам спинного мозга, а следовательно, управляют мышцами вдоха.
2)Каудальная часть ВДГ состоит только из экспираторных нейронов. Все экспираторные нейроны направляют аксоны в спинной мозг. При этом 40%
экспираторных нейронов иннервирует внутренние межреберные мышцы, а 60%
—мышцы брюшной стенки.
Функции нейронов вентральной группы отличаются от функций нейронов
дорсальной группы:
а) во время спокойного дыхания нейроны вентральной группы остаются почти полностью неактивными (выдох совершается под влиянием эластической
тяги легких и грудной клетки);
б) нейроны не участвуют в основной ритмической осцилляции,
регулирующей дыхание;