

1 курс / Психология / Поливагальная_теория,_нейрофизиологические_основы
.pdfП о л и Б А ТА Л Ь Н А Я Т Е О Р И Я
с целью вызова нейрогенной брадикардии, это могло бы привести к катастрофическим последствиям для кислородозависимых органов: коры головного мозга и миокарда. Эта стратегия быстро бы приводи ла кишемии сердца и кортикальной аноксии.Результатом этого была бы смерть. У водных же млекопитающих, наблюдается рефлекс погруже ния, или рефлекс ныряльщика, характеризующийся регулируемой ней рогенной брадикардией, снижающей метаболические потребности. Для выживания, водные млекопитающие выработали, недоступные су хопутным, сложные механизмы управления кислородными ресурсами и смещающие приоритеты в использовании кислорода во время дли тельных периодов погружения в воду.
Возможно, что у млекопитающих во время стресса, когда метаболи ческие потребности велики, а вагусный тонус ДЯ отсутствует, водите ли сердечного ритма (синоатриальный и атриовентрикулярный) мо гут быть склонны к нейрогенной брадикардии, опосредованной ДЯБН. Нейрогенная брадикардия может быть опасной и даже летальной. На пример, при дистрессе плода, когда брадикардия развивается во время гипоксических эпизодов, или она может бы;гь фактором синдрома вне запной детской смерти, или внезапной коронарной смерти у взрослых. В рамках этой модели на собаке было продемонстрировано, что прогрес сирующая асфиксическая гипоксия не только вызывает усиление вагус ного влияния на сердце, но также повышает чувствительность синоатри ального узла к эфферентным воздействиям блуждающего нерва (Potter & McCloskey, 1986). Таким образом, во время гипоксии значительная бра дикардия может поддерживаться при ограниченной или сниженной эф фекторной активностью блуждающего нерва.
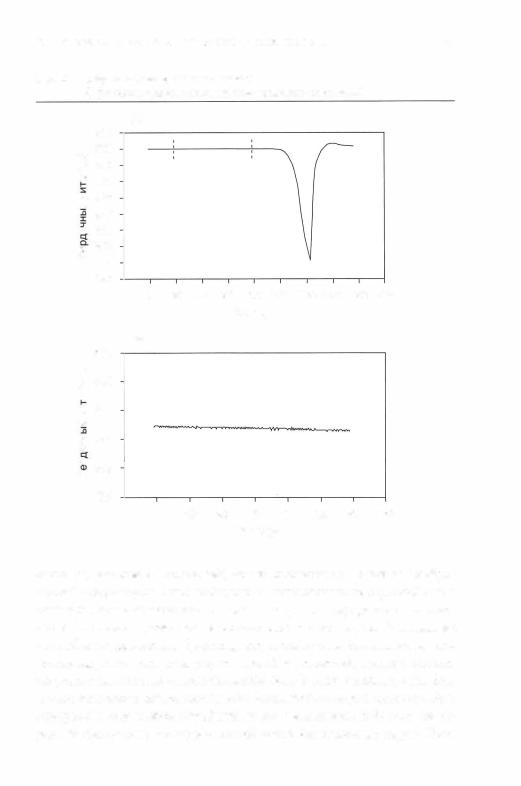
Поливагальная теория дает объяснение выраженной нейрогенной брадикардии, наблюдаемой при дистрессе плода и у новорожденных свысокимриском, у которыхпрактически отсутствует ДСА. Например, какпоказанонарис.2.2,когдавовремя дистрессаплоданаблюдаетсязна чительная брадикардия (рис.2.2а) на фоне низкой вариабельности сер дечного ритма (рис.2.2б).Аналогично и новорожденные с наименьшей выраженностью ДСА подвержены наибольшему риску апноэ и бради кардии (Sostek, Glass, Molina & Porges, 1984).Таким образом, снижение вагальных влияний от ДЯ, ответственные за снижение выраженности ДСА, по-видимому, связаны с повышенной уязвимостью от нейроген ной брадикардии.Поттери Макклоски (Potter& McCloskey, 1986)пред ложили объяснение того, как угнетение функции центральной нервной
61
|
|
|
(а) |
|
|
|
|
|
|
|
|
|
u |
300 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
350 |
|
|
|
|
|
|
|
|
|
|
|
|
|
400 |
|
|
|
|
|
|
|
|
|
|
|
:а |
450 |
|
|
|
|
|
|
|
|
|
|
|
Q. |
500 |
|
|
|
|
|
|
|
|
|
|
|
>S |
|
|
|
|
|
|
|
|
|
|
|
|
|
550 |
|
|
|
|
|
|
|
|
|
|
|
Q) |
600 |
|
|
|
|
|
|
|
|
|
|
|
Q) |
650 |
|
|
|
|
|
|
|
|
|
|
|
() |
700 |
|
|
|
|
|
|
|
|
|
|
|
|
750 |
|
о |
20 |
40 |
60 |
80 |
1 00 |
1 20 |
1 40 |
1 60 |
1 80 |
|
|
|||||||||||
|
|
|
||||||||||
|
|
|
|
|
|
|
Секунды |
|
|
|
|
|
|
300 |
|
(б) |
|
|
|
|
|
|
|
|
|
u |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
320 |
|
|
|
|
|
|
|
|
|
|
|
:а |
|
|
|
|
|
|
|
|
|
|
|
|
5. |
340 |
|
|
|
|
|
|
|
|
|
|
|
>S |
|
|
|
|
|
|
|
|
|
|
|
|
|
360 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
Q) |
|
|
|
|
|
|
|
|
|
|
|
|
()Q. |
380 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
400 |
|
о |
1 0 |
|
20 |
30 |
40 |
|
50 |
60 |
70 |
|
|
|
|
|||||||||
|
|
|
|
|
||||||||
Секунды
Рекомендовано к покупке и изучению сайтом МедУнивер - https://meduniver.com/

Поттер и Макклоски в этой работе не исследовали дел, мы можем предполагать, что делв их опытах не была выражена, посколькуживот ные были анестезированы до хирургических, электрических и гипокси ческих манипуляций, и поскольку гипоксия и анестезия связаны с пода вленной вариабельностью сердечного ритма, включая дел (например, Donchinet al., 1985; Nelson, 1976).
Возможность бифуркации вагальных влияний так же демонстрирует электростимуляция ДЯБН у кролика. Как показано на рис. 2.3, электри ческая стимуляция ДЯБН приводит к брадикардии без увеличения дел. Это контрастирует с эффектом депрессорного нерва аорты, который свя зывается как с ДЯ, так и с ДЯБН. рис. 2.4показывает, что у кролика, нахо дящегося под таким же наркозом, стимуляция депрессорного нерва аор ты приводит к увеличению дел и выраженной брадикардией (например,
МсеаЬе, Yongue, Porges & & Лckles, 1984).
Поливагальная теория утверждает, что волокна блуждающего нерва, идущие от ДЯБН и ДЯ отличаются по структуре и функции. В частно сти, уже говорилось, что вагусные эфферентные волокна из ДЯ миели низированы и угнетают дыхательный ритм, а эфферентные волокна, иду щие из ДЯБН, немиелинизированы и не влияют на дыхательный ритм. Однако существуют наблюдения, противоречащие описанному ранее.
lэые4й4й 5н ли0внлиушlчу вжжву г |
е0тылмгиoЕ уциен |
ь1 I з в,емтеиинорв,,оу м |
|
лл |
н |
н |
аж |
0нлни0вЭ
ппкс
покс
тp пйкс
гр
выp пюкс
гр чккс
чпкс
чокс
цс пкс окс йкс юксеккс е пкс е окс мрeи.лс

63
220
u 240
';' 260
f
:s:
· 280
I :т
ф
300
uф
320 |
|
|
|
|
|
|
340 |
20 |
|
|
80 |
1 00 |
1 20 |
о |
40 |
60 |
Секунды
Например, Д. Джордан с соавт. (Jordan et al., 1982) сообщали, что су ществуют кардиоингибирующие вавгусные нейроны, происходящие из ДЯБН, с эфферентными аксонами, проводящими в диапазоне В-воло кон, и, следовательно, миелинизированные.Более того, указанные авторы утверждали, что эти нейроны угнетают дыхательный ритм.Хотя данные полученные Джорданом с соавт., подтверждают предполагаемый двойной источник вагусных эфферентов, их выводы противоречат предлагаемому функциональному различию.
Существует несколько возможных объяснений несоответствия, вы явленного Джорданом с соавт. Первым может быть метод. В своем ис следовании Джордан и соавт. использовали стандартные нейрофизио логические методы стимуляции и записи для идентификации нейронов. Согласно Дж. Шваберу (Schwaber, 1986), многие волокна блуждающего нерва, ранее предположительно происходящие из ДЯБН, были иденти фицированы спомощью более новыхметодов, такихкак ретроградно ме ченная пероксидаза хрена, как исходящие из ДЯ. Он также заявил, что, поскольку аксоны из ДЯ проходят очень близко к границе ДЯБН, труд но стимулировать или повреждать ДЯБН не затрагивая ДЯ, искажая ре зультаты исследований по электрической стимуляции. Таким образом, дополнительные исследования с более точными методами маркировки могут продемонстрировать, что все нейроны, проводящие в диапазоне
Рекомендовано к покупке и изучению сайтом МедУнивер - https://meduniver.com/

|
|
|
|
|
|
|
|
|
|
|
|
П о л и В А Г А А Ь Н А Я 1' Е О Р И Я |
|||||||
|
."'1"'/"'ЛО'.Б"'121 ' |
l |
' |
' 1'ЛЛ1'О1"'1 |
В'ЛЛ1'. "'111'Б Л ' /'"'1'о |
||||||||||||||
/'БО1''1. 1'.1'ЛЛ1' Л' ''.Т |
Т' |
l |
О1"'11.''Б1'Б 1 ."' ."'БЛ"'.1АЛ'1''о |
||||||||||||||||
.'1АЛ "'y,.''/''"'./'l |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
"'.1"'.'. |
Б"''1'Л' . ."''.1l Я |
Б Д Г ' lО |
|
..лО' Л'y,'"'Л1О."' |
||||||||||||||
А2Б1''.1' |
.1'. |
2И''y,1'1''.."''Л1Б |
Л''."'. |
'"''11О 2ГЛ'1'1' |
11Б'1'1 |
||||||||||||||
Л1y,''1 |
.Г Л'1"'Г'ЙЛ"'/'/О |
1"'/' |
'Л"'Л' ''.2Л/' |
|
l |
О.1'. 2И'21'1''.о |
|||||||||||||
."''Л"'Г"''"''1'1АЛ"'Л"'Л''.' |
.1 |
1.'1' |
/'/ А''1'/'' |
1'БО1'/ |
' 2.'1'Й 'Л'' |
||||||||||||||
|
l |
1Л'/"'.1'. |
оИ'2 |
|
|
.1 |
1.'1' |
1"'1А/"'Л' ЛЙ' '1'1АЛ2БА''о |
|||||||||||
1'/''1'Б |
|
1' 1'ЛЛ1' |
..'1'1'1А.1.2Б1 |
"' 1"'.ОЙ1"'.'Г2. Л'2 '.1 |
|
.' |
|||||||||||||
И'2 1"'.1' |
.1'. |
2И'' |
|
|
Л' |
.1 |
2'1 |
Л' |
11Б'1'1АЛ1y, ''1 .Т |
"'1'' |
|||||||||
1"'Г"'ОА''1'/''1'2 А11' |
Л'.'11'ЛЛ"'y,' |
1"' |
1'1' |
Л1Л"'.1' Л'1"'.'Л '1' |
А'' |
||||||||||||||
1'/''1'БО ."' |
Л'/'БИ2Б. "'1.'1 Л'.1'.212И'Б |
1'1''.."''Л"'Г"' |
'"''1 |
' |
|||||||||||||||
Л"'Г"'Л''.' |
'1'Й'Л' А''1'/'' |
1'' |
."'.1 |
'. |
21' |
"'/"'1"' |
"'1 .'1'Й'Л 1О |
||||||||||||
.1 |
.'ЛЛ"'y,.1'. 12И''y, '"''1'1АЛ"'Г"'Л''.'О .'1"'1"'.О 1''11"'1'Г' |
И'. |
|||||||||||||||||
''/'21''"'.'Л'' |
.'Г 2.Л1Б ."'1"'/"'Л /'/ |
' |
|
О1'/ |
' |
' |
|
|
l ''1' |
||||||||||
.1.'Л'' |
|
1 1'1'.21'1'"'И'Л1Л"'Л"' |
.1'2Л'2 |
/'Б1"'y, .'Г'1АЛ"'y,.'. 1'.' |
|||||||||||||||
Л' .''1' |
ЙЛ1y,''1 . |
1'' |
А''"'''Н1'/.' |
."' Г1'.2 '1.2 . |
1'ЛЛ1.' |
Г 'Й'1"' |
|||||||||||||
' |
l '"'1' Я |
Д Д Б |
"' |
|
О л'2lЛ'2."' ."'БЛ"'.1А."'.1"''1 |
. 1"'.О |
|||||||||||||
Й1"'.2И'.1.2Б1 .'1"'.1' |
'' |
1'Й'2 . |
"''Л'Л' |
'И'' ' |
Н2Л/И'"'Л''"'.'Л'' |
||||||||||||||
|
|
|
Т'1' '.''О |
11о"'А1'ГЙ'Л'о'.."'А'1' |
'И''О |
2 /'"'1'/' |
."'Л ' '' |
||||||||||||
.'1 |
А.2 2Л'/'1 |
Л1' .'Л2.Л1' .''1'Л"'.1' |
|
121' |
"'1 |
|
О/"'1"''1' |
Л' ' |
|||||||||||
.'.21 "'1 11Б'1'1АЛ"'y,Н2Л/И''Т "'Г1'.Л"' уч1"'."'2А |
2.Л'Л'БО |
|
А2 |
||||||||||||||||
1'1 '.'1А |
|
|
."'1"'/ЛОЛ"'"'Л' Л' А2121.1 |
21АЛ' 11Б'1'1АЛ1y,''1 . |
|
'"'.' |
|||||||||||||
1"'Г"'2ОЛ'/"'1"''1Б .'1"'. |
.1'/"'1'1 |
'БИ'Б. .'."'. |
|
'1' |
.А1' |
' Л'Л"' |
|||||||||||||
."'Г21 ''.1"'1'Л'1А.2 |
Л'y,'"'Л1О2. |
оБИ''.2 |
Й'.1АБ"'АИ'Г"'/''1'"'1 |
о |
|||||||||||||||
."'Л''Л"'Г"'."'1'1 '12 ''1 |
.' |
|
|
|
ГР |
ГРО лl |
|
|
|
|
|||||||||
|
'1 |
|
Л'y, |
''..1'1"'.'Л'2 |
|
1"' ."'1о1"'1''1'1'1АО |
Л'./"'1А/"' |
1"'ЙЛ1 |
|||||||||||
1''11"'Б'ЛЛ1' |
Н2Л/И'"'Л'1 Л1' |
' .1'2/1 |
2'Л1' |
'' |
1'Й'2 .'Б12 |
учН |
|||||||||||||
Н'''Л1'.' |
|
|
' |
О.Н"''.21''"'.'Л |
|
Л1' |
1"'1 |
.'Л'1АЛ"'y,1'"''''y, |
|||||||||||
|
И' "'1Л' 1'"'А1'.' ..2 |
'Л' . |
"'А"'АИ'Л''. |
'' |
|
|
А1'1"'Л.' ..'Б .1'/"'о |
||||||||||||
1'1'БИ'БТ |
"'1А '2Й'.1АЛ'y,'"'Н' |
'"'1"'Л'Й'./ |
Б' Л'y,'"''Л'1"'.'Й'о |
||||||||||||||||
./'Б '..1'1"'.'Л'y, |
А12Б1'БИ'БЛ''."'. |
2 .1'/"'1'1' |
|
БИ'Б А11' |
1'"'.'о |
||||||||||||||
1'Л' Л' /'1.'БО /'"'1'/'БО /"' |
/'Б' ."'А'/'Бl |
..1'1"'.'Л'2О ' |
2В'БИ'' |
||||||||||||||||
.'Л2.Л2Б ''Г |
2И'Б2 1 |
1'y,О"'Г''Л'Й'.' |
'.А '..1'1"'.'Л' |
|
.' |
Н''.' |
|||||||||||||
/"'1"'Г'Й'./"'y, А1"'/'11 |
. 1'''Н'' |
|
'Й'./'.' |
Н' '"'1"'Г'Й'./'.' |
' |
.''' |
|||||||||||||
Л'2.'l |
|
' 1'/ |
2Б .Л"'Л"'Л'y,'"''Л1"'. 'Й'./'Б '..1'1"'.'Л'y, |
.1."'1' |
."' |
||||||||||||||
Г' Й1"'.'/' 1'"'."'1'1.2 |
Л' 1'И''Л1 'БО2.'' |
'Б "'1 А"'1' |
Л'y, ' |
1' '..l |
|||||||||||||||
|
'/'. "'А'' "'.О ."'БЛ"' 2."'.Л'1А.2 |
. |
"'А"'АИ''."'.1' 1"'1'.'Г'1 |
|
Л"'y, |
||||||||||||||

М И Р Е , , , |
65 |
модели, разработанной на основе исследования ствола мозга грызунов для человека. Однако существующие данные иллюстрируют такие яв ления, как клиническая брадикардия при отсутствии ДСА у плода че ловека (см.рис. 2.2), сдвиги ДСА, не зависящие от изменения частоты сердечных сокращений во время ингаляционной анестезии (например, Doncl1in et al., 1985) и короткие задержки реакций от обеих систем (см. рис. 2.3 и 2.4), которые подтверждают аргументы в пользу поливагаль ной модели.
Ваrальные стратегии у млекопитающих и рептилий
Вагальные системы рептилий и млекопитающихвыказывают противопо ложные стратегии. Для рептилий характерен низкий общий ваrальный тонус и его кратковременное повышение в ответ на вызовы окружаю щей среды. Напротив, для млекопитающих характерен высокий общий вагальный тонус и его кратковременное снижение в ответ на средовые вызовы (табл.2.3).
Таблица 2.3. Вагал1,ные стратегии
Поведенческие стратегии рептилий способствуют их выживанию и приспособлению к окружающей среде. Большинство поведенческих паттернов связаны с поиском пищи, преследованием и едой.Только не значительные временньrе и энергетические ресурсы направлены на со циальные взаимодействия, такие как размножение и воспитание потом ства. В оборонительной стратегии рептилий нейрогенная брадикардия является адаптивной реакцией и не ставит под угрозу их физиологичес кий статус.Урептилий, в сравнении с млекопитающими, органы тела ме таболически менее активны, имеют отличные метаболические механиз мы и менее зависимы от кислорода. Они даже способны долгое время обходиться без кислорода.Адаптивная стратегия рептилий смертельна для млекопитающих. Оборонительная стратегия млекопитающих тре бует ускоренного метаболизма, способствующего реакции «сражай ся или беги». Следовательно, рефлекторная длительная нейрогенная
Рекомендовано к покупке и изучению сайтом МедУнивер - https://meduniver.com/

брадикардия будет уменьшать кислородные ресурсы и метаболический выход и ставить под угрозу потенциал борьбы или бегства млекопитаю щих. Последствия сокращения кислородных ресурсов также могут уг нетать функции центральной нервной системы, снижать сложность по ведения и правильное выполнение сложных поведенческих паттернов, вызвать бессознательное состояние, вредить жизненно важным органам и, в конечном итоге, при достаточной длительности, привести к смер ти.Таким образом, кардиальный компонент ориентировочного рефлек са должен быть кратковременным и заменяться физиологической реак цией, которая не нарушает кислородную потребность нервной системы млекопитающих. Этой цели служит снижение вагусного тонуса, возни кающего в ДЯ.
мггьпнпэм, пльмпл лкгьмлблэкпшнгелелиллндeлшпльсм2л
Нейрогенная брадикардия, регулируемая ДЯБН, которая наблюдает ся у рептилий и млекопитающих во время ориентировочного рефлекса, могла развиться из системы вкусовых рефлексов примитивных позво ночных. Густация является основным методом идентификации добы чи (включая другие соответствующие источники пищи) и хищников в воднойсреде.Например, у рыб недифференцированная вагусная доля контролирует вкусовые, алиментарные и пищеварительные процессы (Finger & D1111widdie, 1992)смpРефлекторное повышение тонуса блужда ющего нерва затронуло бы несколько органов: сердце, где оно умень шило бы обмен веществ и дало бы возможность животному мгновенно застыть; органы,содержащиевкусовые рецепторы, гдеоно сориентиро валось бы на источник стимуляции и регулировало порог обнаружения новизны; и пищеварительную систему, где оно стимулировало бы желу дочную секрецию и моторику.
В процессе филогенетического развития висцеротропная организа ция системы блуждающего нерва усложнилась благодаря включению путей других черепно-мозговых нервов, включая тройничный, лице вой, добавочный и языкоглоточный. Таким образом, в вагальную си стему интегрировались более специализированные функции, такие
мргэондвцпу ог разиорс. няцябгонорлв sорян лоигигятв п р но(орон sотоигинсо бнояцпнзягиуранзблся цял(нс Млллянрвлтицярсялотп и л лянрвпрвoзилсялояупi пли пдявг ито,лсн лянноллсн иои(жри итоии(зв лло nNитояр(игнза(знзЭпотвигМ бняли(вригятигрв рлзгняллп. ьядоня1ябгонор з нсо (обоoнвэпияиап ля оглятялв олразиорс. аол лрвDзилвулоту(Э

О Б О Р О Н И Т Е Л Ь Н О М |
67 |
как вращение головы для ориентации сенсорных рецепторов в направ лении источника стимуляции, жевание для поглощения пищи и слюно отделение для инициирования вкусовых и пищеварительных процессов.
Моторный компонент блуждающего нерва имеет эволюционное происхождение от четырех черепно-мозговых нервов (тройничного, лицевого, добавочного и языкоглоточного). Блуждающий нерв не толь ко иннервирует гладкую мускулатуру и миокард, но, подобно другим четырем черепно-мозговым нервам, содержит двигательные пути, ин нервирующие поперечно-полосатые мышцы. Вагусные пути, иннерви рующие соматические мышцы, часто не включают в нейрофизиологию вегетативной нервной системы. Эти волокна называют
чтобы отличать их от двигательных путей, иннервирующих гладкие мышцы
и миокард. Основное различие между двумя типами двигательных пу тей заключается в том, что соматическая регуляция мышц может быть сознательной и произвольной, тогда как регуляция гладких мышц реф лекторнаи неосознаваема.Поскольку специальные висцеральные эффе ренты иннервируют произвольные мышцы, их, как правило, исключают из вегетативной нервной системы. Традиционно, только общие висце ральные эфференты как из симпатической, так и из парасимпатической ветвей относят к вегетативной нервной системе.
Соматические мышцы, иннервируемые пятью черепно-мозговыми нервами, формируются из висцеральных дуг, эмбриологически извест ных как жаберные дуги (Warwick & Williaшs, 1975). Эти мышцы име ют решающее значение для поведения некоторых видов млекопитаю щих. Например, соматические мьшщы, иннервируемые тройничным нервом, развивающиеся из первой жаберной дуги, участвуют в жева нии, втягивании нижней челюсти и закрытии рта. Специальные висце ральные эфференты лицевого нерва, возникающие из второй жаберной дуги, иннервируют мышцылица, кожиголовы и шеи, обеспечивая мими ку. Лицевой нервиннервирует также мышцы нижней части рта.
Тройничный и лицевой нервы развиваются из жаберных дуг и име ют связь с тремя другими черепно-мозговыми нервами, также проис ходящими из жаберных дуг, исходные ядра специальных висцеральных эфферентов языкоглоточного, блуждающего и добавочного нервов ис ходят из того же ядра - ДЯ. Таким образом, эфферентные волокна проходят через три разных черепных нерва, но берут начало в одном
и том же ядре.
Рекомендовано к покупке и изучению сайтом МедУнивер - https://meduniver.com/

|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Т Е О Р И Я |
|
|
|
У 2''yv"'.."'Б'2'1"''"'''Б"' |
.l' |
' '.D1'''1 |
'.А'1'3"' |
11'. |
.2"'yv'.А'3А |
|
||||||||||||
|
|
|
|
|
|
|
|
|
|
Ф |
|
|
|
|
|
|
|
|
|
1'.yv"'.2А'3А ПББ'"''''3А |
2В'"'Б 1 1D3l'12'''Б''1А |
А2ВБ1.БВ"'1 |
' 1' |
|
|||||||||||||||
А.1'Б''1 |
'"''1.А 1'1'''ВБ' |
. |
'А'.D'1.''"'1 |
ЛЯЙ |
"'l'2''.БВ'А |
ЛЯ |
|
||||||||||||
В2'.121"'' |
.2'Б |
'Бl'''1''.yv'"'Б 12 ''l'А 111l'1' |
'"'А.А1'''.'' |
' 2'В"'Г |
|
||||||||||||||
1'1.Н У.'А'"' D'.Б"'''"' |
121 |
2.'А'Б'D ''2'1'Б"'.l'А |
2''yv"'..'1 '1"'"'' ''А |
|
|||||||||||||||
Б'' ''"''А1 |
Б.А"'''.1 |
1 |
1. |
'.АБ"' 1."'' '.Б.2' |
.''''Б |
.''"''''А |
.'1"''Б.Г |
|
|||||||||||
В"'Б2"'''Б"'''Б"'.l'"' А"'1''"'yv"'2'''3АБВ1.'1''"'2А'3"' |
l В''1'11 |
А'.2'Г |
|
||||||||||||||||
''1. |
' |
В12"'l'.2'1' |
1.D. ЛЫЕ |
РЫСС |
ЬЬЕункцТАТСоНН''1"' |
''1'А 1'А.1'БЕ |
|
||||||||||||
'3Б '"''1 |
.'1"''БЦ' 1'2'l'.А А"'ВВ'"' '.Б.2' |
1 О"'Б''1 |
''1"'2"' .2''''1' |
|
|||||||||||||||
1'D1. |
l''''3"'А |
'''"''1''ВБ' |
|
13Оyv3А''1"'Б.БВ'"' D. 2'2'Б"'''"' |
' |
11'Е |
|
||||||||||||
Б"'''1 |
"''В УАВ'"' .'''3"' |
.''"''''А 1'В''"''''"' |
1'"'1'3"' |
1"''3 ' |
А2ВБ1.Е |
|
|||||||||||||
БВ'"' '"''13 2''А'11' |
|
1 12ВА''"' П''А13ОyvЛЫЕЮ |
Р ЫСЬЬ ЕункцТАоС НЙ |
||||||||||||||||
|
.l'1 |
'А'.D'1А П''' |
l'12И"'l. |
'А2.1."'' |
.2'.'А''.'АБ |
2' |
.'"'1.'1'1 |
.'Г |
|
||||||||||
1.''Б"'.А'А 13Оyv'''"''''''1 |
|
.'А 1'. yv"''.2А'3"''"'yv"'2'''3А l'''1' |
'''' |
|
|||||||||||||||
1.'А .''Вl'В'3А .11D.''3"' |
. 12 ''.''"'1 |
|
' '''31'1.''"'1А |
' '"'1В2'''1.'А |
|
||||||||||||||
1'1'lВ |
' П1'yv''Й'' |
|
11' 1.'"'2А'3"' |
11'. |
2' |
Б.Б' ''Б ''1.yv'Б '' |
l''3 |
|
|||||||||||
1'2'1''1' |
|
1'D1. |
121 |
l'''1''.yv'' |
.2'Б''1' |
l'122"'l.''1' |
2'1 "'1"'''1 |
|
|||||||||||
. ."''1"'Б''лу2"'1'Б''Б БВ'lyv'"'БЙуюl'1. |
'А '.D'1А |
Б'2'1"''"'''Б"' .l'А 1.Б"' |
|
||||||||||||||||
l'11. |
Б.А"'''3"' |
1В 1' |
''.'.Б'' |
1''ВБ'.1 |
1 А'.'АЦ'1"'''3"' |
13Оyv3, |
|
||||||||||||
|
|
, |
|
, |
, |
|
|
|
|
|
|
, |
|
|
, |
|
|
|
|
|
|
|
|
|
|
|
, |
|
|
|
|
|
, |
|
|
|
|
||
|
|
|
|
|
|
|
|
|
, |
|
|
, |
|
|
|
|
, |
|
|
|
|
|
, |
, |
|
|
|
|
, |
|
|
|
|
|
|
|
, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
, |
|
|
|
|
|
, |
|
, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
, |
|
|
|
|
,, |
|
|
|
|
|
, |
|
|
, , |
|
|
|
|
, |
|
, |
|
, |
|
|||||
|
, |
|
|
|
|
|
|
|
|
|
|
, |
, |
|
|||||
|
|
|
|
|
|
|
|
|
|
|
, |
|
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
, , , |
, |
|
, |
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||
|
|
, |
|
,, |
|
|
, |
|
|
, |
|
|
, |
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|||||||||
|
|
, , |
,, , , , |
|
, , ,, |
|
|
, , , , |
|
||||||||||
|
|
|
|
, , , |
|
|
, |
|
|
|
|
|
|
|
|
, |
|
|
|
, |
|
, |
|
, |
|
|
|
|
|
|
|
, |
, |
|
|
, |
|
, |
, |
|
|
|
|
|
|
|
|
|
|
||||||||||
|
|
, |
|
|
|
|
|
|
|
|
|
|
|
|
|
,, |
|
|
|
|
|
, |
|
, |
|
|
|
|
|
|
|
, |
|
, |
|
|
|
|
|
|
|
|
|
, , , |
, , |
|
, |
|
, , , |
|
, , , |
|
|
|
|
|
|||
|
|
, |
|
|
, , , |
|
|
|
, |
|
|
|
|||||||
|
|
, |
|
|
|
|
|
,, , |
|
|
, |
, |
, , |
|
|||||
|
|
, |
|
|
|
|
, |
, |
|
|
,, |
|
|
|
|
|
|
|
|
|
|
|
, |
|
|
|
|
|
|
|
, |
, , |
, |
|
|
|
|
||
|
|
|
, |
, |
|
|
, |
, |
|
|
, |
|
|
|
|
|
|
|
|

69
интонацию вокализацией. Таким образом, эфферентные проекции ДЯ управляют процессами, связанными не только с питанием и дыханием, но также с движением, эмоциями и общением. Такая организация спо собствует уникальным стратегиям выживания и социальному поведению, которые наблюдаются у млекопитающих. ДЯ осуществляет вагальное торможение, которое млекопитающие мгновенно устраняют для увели чения метаболического выхода, способствующего реакции «сражайся или беги». В вагусном ДЯ берутначало двигательные пути, обеспечиваю щие интонирование вокализаций (например, паттерны крика)для прояв ления эмоций и передачи внутренних состояний в социальном контексте.
Поведенческие дериваты двух ветвей блуждающего тела предполага ют типологию, в которой одна ветвь имеет дело с бессознательными реф лекторными вегетативными функциями, а другая участвует в более со знательной, произвольной, гибкой и часто социальной деятельности. Эти типологии можно обосновать и с точки зрения нейроанатомии. ДЯБН содержит только общие висцеральные эфференты, которые иннервируют гладкие и кардиальные мышечные волокна и управляют секрецией желез. В то время, как в ДЯ локализованы специальные висцеральные эфферен ты, которые иннервируют произвольные поперечно-полосатые мышцы мягкого неба, гортани, глотки и пищевода.
Связь соматомоторнойи висцеромоторной систем У млекопитающих мы наблюдаем две эволюционные стратегии, которые
связывают вегетативные функции с соматической мышечной активно стью. Во-первых, существует анатомическая связь между сегментацией спинномозговых нервов и симпатическими цепями. Эта связь отражает ся в связанном с движением повышении симпатического тонуса, кото рое преследует психофизиологов, стирая грань между моторными и ве гетативными реакциями. Эволюция сегментированной симпатической нервной системы шла параллельно эволюции произвольной двигатель ной активности. Симпатическая нервная система регулирует вазомо торный тонус, направляя кровоток, а соответственно, и кислород к кон кретным мышцам, подвергающимся стимуляции. Кроме того, существует симпатическая судомоторная иннервация, которая регулирует потоотде ление, увлажняющее кожу и защищающее ее от разрывов. Эта связь меж ду симпатической активностью и движением была краеугольным камнем теории возбуждения и гипотез, связывающих вегетативную функцию
Рекомендовано к покупке и изучению сайтом МедУнивер - https://meduniver.com/