
Биосинтез липидов
.pdfБиосинтез липидов
Биосинтез липидову животных представляетсобойоченьактивный метаболическийпроцесс.Взначительнойстепениэтообусловленнотем,что животные могутзапасатьэти соединенияв большихколичествах.Надолю триглицеридовворганизме мужчинывесомв70кг приходится около12кг.Втех случаях,когдауглеводыпотребляютсяс пищейвизбытке иорганизмне способен откладывать ввиде гликогена,онипревращаютсявтриглицериды,которые накапливаютсявклетках жировойткани. Больше всеготакихклетокнаходится в подкожнойжировойклетчатке ивбрюшнойполости. Растениятакже могут запасатьэнергию ввиде высококалорийноготоплива –триглицеридов,особенно многоэтихсоединений вразличныхплодах,орехахисеменах.
Полярные липидымембран (фосфолипиды исфинголипиды)ворганизме животных не запасаются,ноонитоже постоянносинтезируются длявосполненияпотерь, обусловленныхразрушениеммембранвходе метаболических процессов.
Биосинтез жирныхкислот |
|
|
Жирныекислоты являются основнымистроительнымиблоками триглицеридови |
||
полярныхлипидов. |
|
|
Значительнаячастьзаменимых жирныхкислотсинтезируетсяв печени,вменьшей |
||
степени — вжировой тканиилактирующеймолочнойжелезе. Вэукариотахсинтез |
||
жирныхкислотпроисходитв |
|
всостоянии покояилипосле еды. |
Условноможновыделить4этапабиосинтеза: |
||
|
цитоплазме, |
|
1. Образование аце ил-КоА из глюкозы,другихмоносахаров или кетогенных |
|
ами окислот.Избытокуглеводов, поступающихворганизм,трансформируетсяв |
|
2. Перенос ацетил-КоА из митохондрий в ц тозоль |
|
жирные кислоты,азатемвжиры. |
|
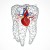
Ацетил –КoA образуетсявмитохондрияхиз пирувата(:в пируватдегидрогеназном |
|
комплексе),из жирныхкислот ипри расщепленииаминокислот, а все последующие |
|
реакциисинтезажирных кислот происходятвцитоплазме. Какмыуже знаем |
|
митохондриальная мембрананепроницаемадляацетил-КоА.Какимобразом |
|
|
помощи челночного механизмапереносаацильных |
попадаетмитохондриальнаяацетил-КоА вцитоплазму? |
|
Этапроблемарешаетсяпри |
|
Вначале внутримитохондриальный ацетил-КоА взаимодействует соксалоацетатом, |
||||
групп через тох ндриальную мембрану. |
|
|
|
|
врезультате чегообразуется→цитрат (I реакцияциклаКребса) . |
||||
Ацетил-КоА +Оксалоацетат Цитрат+HS-КоА. |
|
.Образовавшийсяцитрат |
||
Реакциякатализируетсяферментом |
|
|
|
|
переноситсячерезмембранумитохондрий вцитозольпри помощиспециальной |
||||
|
цитрат-с |
нтазой |
|
|
трикарбоксилат-транспортирующей системы |
. |
|
||
|
|
|||
Вцитозоле цитратреагируетсHS-KoA иАТФ,вновь распадаясьнаацетил-КоА и |
||||
оксалоацетат. |
→ |
Ацетил-КоА +АДФ+Pi+ Оксалоацетат |
||
Цитрат+HSKoA +АТФ |
|
|||
ЭтареакциякатализируетсяАТФ- |
|
.Ацетил-КоА вцитоплазме служит |
||
исходнымсубстратом |
|
|
цитрат-лиазой |
|
длясинтезажирных кислот,аоксалоацетат уже вцитозоле, при участиицитозольной малатдегидрогеназы восстанавливается до малата.Последний при помощи
дикарбоксилат-
системытранспортирующвозвращаетсявй митохондриальный матрикс,где окисляется дооксалоацетата, завершаятемсамымтак называемыйчелночный
цикл.Надо отметить
что перенос цитратав
цитоплазму происходит только ввидецитрата
при увеличении количествацитрата
вмитохондриях, когдаизоцитратдегидрогеназапериодеα- кетоглутаратдегидрогеназа.Этаситуациясоздаетсявингибированыабсорбтивном выс кими,когдаконцклеткантрациямипечениполучаетНАДНи
достаточноеАТФ количествоисточниковэнергии.
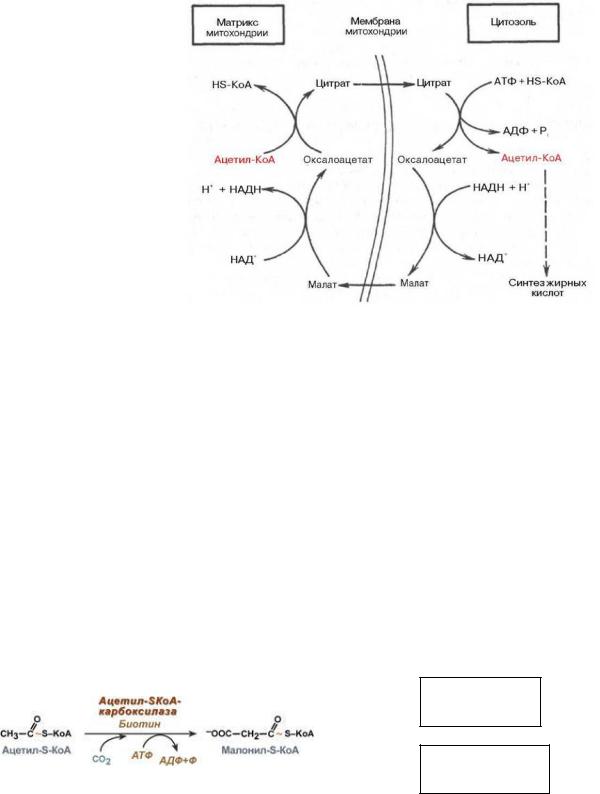
3. Образование малонил-КоА из ацетил-КоА.
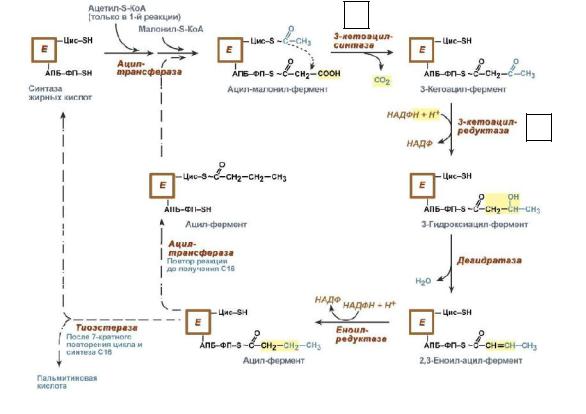
является первой реакцией синтеза жирных кислот. Малонил-КoA синтезируется из ацетил-КoA и двуокиси углерода (СО2). Фермент, катализирующий эту реакцию - ацетил-КоА-карбоксилаза, очень сложный фермент. Он содержит ковалентно связанный биотин. (Биотин является водорастворимым витамином группы В. С участием биотина протекают реакции активирования и переноса СО2). В первой стадии реакции С02 ковалентно связывается с биотином, во второй стадии СО2 группа переносится на ацетил-КоА с образованием малонил-КоА. (Биотиновя группа служит подвижной рукой, переносящей СО2 на ацетил-КоА.) Присоединение новой карбоксильной группы к ацетил-СоА обеспечивается за счет энергии АТФ. Ацетил- КoA-карбоксилаза - регуляторный фермент, который определяет скорость всего процесса биосинтеза жирныхкислот вживотныхтканях.
ацетатCH3-COOH
малонатCOOH-CH2-COOH
Биосинтез жирных кислот катализируется |
|
|
|
. |
Синтазная |
|||
4. Синтез пальмитиновой кислоты. |
синтазой жирных кислот |
|
|
|||||
система для жирных кислот катализирует суммарную реакцию, в ходе которой одна |
||||||||
молекула ацетил-КоА и семь трехуглеродных молекул малоновой кислоты в виде |
||||||||
тиоэфира малонил-КоА последовательно соединяются друг с другом, в результате |
||||||||
чего образуется 16-углеродная пальмитиновая кислота и выделяется семь молекул |
||||||||
СО . |
|
|
|
|
→ |
|
|
|
2 |
|
|
|
|
|
|
|
|
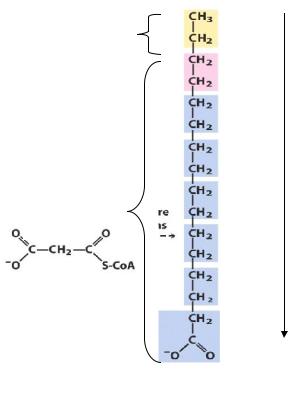
ацетил-КoA + 7 малонил-CoA + 14 НАДФН + 20 H+ |
|
|
|
|
||||
|
|
|
→CH3(CH2)14COOH + 7CO2+ 8 КoA-SH + 14 NADP+ + 6 H2O |
|||||
Единственная молекула ацетил-КoA служит «затравкой», углеродные атомы |
||||||||
метильной и карбоксильной групп этой молекулы занимают соответственно 16 и 15 |
||||||||
положение в образовавшейся молекуле пальмитиновой кислоты. Биосинтез жирных |
||||||||
кислот |
начинается |
с ацетильного остатка и |
рост цепи продолжается по |
|||||
направлению к карбоксильномуконцу. |
|
|
|
|
16 |
|||
Источники атомовуглерода |
CH3-CO-S-CoA |
|
15 |
|||||
|
14 |
|||||||
|
|
|
|
|||||
при биосинтезе жирныхкислот: |
|
|
|
|
||||
|
|
|
|
13 |
||||
|
|
|
|
12 |
||||
16и15углеродные атомыберутся |
|
|
|
|
11 |
|||
|
|
|
|
10 |
||||
изацетил-КоА,остальные из |
|
|
|
|
9 |
|||
малонил-КоА. |
|
|
|
|
|
8 |
||
|
|
|
|
|
7 |
|||
Впроцессе ростацепи 1-ыйи 2-й |
3 |
2 |
1 |
|
6 |
|||
углеродные атомымалонильных |
|
|
|
|
5 |
|||
|
|
|
|
4 |
||||
группвстраиваются в цепь,а 3-ий |
|
|
|
|
3 |
|||
атомвыделяетсяв виде СО2 |
|
|
|
|
2 |
|||
|
тазажирныхкислот,осуществляющая |
|
|
|
|
1 |
||
С |
|
|
|
|
|
|||
синтез |
кислот, |
является мультиферментным комплексом, которая состоит |
||||||
из 7 |
индивидуальных ферментов. Центральное местовэтомкомплексе |
|
|
|||||
занимает ацилпереносящий белок (АПБ). С ацил-переносящим белком ковалентно |
||||||||
связывается простетическая группа, которая состоит из фосфопантотеиновой |
||||||||
кислоты и сульфгидрильной группы (SН-группы), подобно SН-КоА. |
Один их |
|||||||
ферментов комплекса, |
3-кетоацил-синтаза, также |
имеет SН-группу |
|
в составе |
||||
цистеина. Взаимодействие этих групп обусловливает начало и продолжение биосинтеза жирной кислоты, а именно пальмитиновой кислоты. Для реакций синтезанеобходимНАДФН.
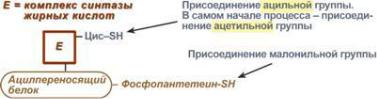
В первых двух реакциях последовательно присоединяются малонил-КоА к |
|||||||||
фосфопантетеину ацил-переносящего белка и ацетил-КоА к цистеину 3- |
|||||||||
кетоацилсинтазы. Важно,что |
После того как обе сульфгидрильные группы |
||||||||
|
|
|
происходит |
|
|
||||
нагружены, |
|
|
|
Малонильная группа связывается только с SН- |
|||||
|
наращивание цепи жирной кислоты - комплекс готов к |
||||||||
группой фосфопант те |
|
. |
|
|
|
|
|||
первому циклу синтеза. Один цикл работы синтазы жирных кислот состоит из 4 |
|||||||||
1. |
Ацетильная группа |
|
|
|
|
с остатком малонила по месту |
|||
отделившегося С02. (переносится с SН-группы цистеина на малонильную группу, |
|||||||||
стадий: |
|
|
конден ируется |
|
|
||||
которая связана с SН-группой фосфопантеина. В ходе этой реакции образуется та |
|||||||||
самаяСО2,котораяисходно включиласьвмалонил-СоА). |
по карбонильной группе |
||||||||
2. |
Полученный радикал ацетоацетила |
|
|
||||||
кетоацил-редуктазой собразованиемгидроксиацил-S-АПБ. |
|
||||||||
4. |
Опять |
|
|
|
|
|
восстан вливается |
|
|
|
|
|
еноил-редуктазой. |
|
|||||
3. |
Дегидратируется |
|
|
|
|
|
|
||
|
восстанавливается |
|
|
|
|
|
|||
1
2
|
|
5 |
|
|
|
|
|
|
3 |
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
6 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
4 |
|
|
|
|
|
|
|
|
|
Для восстановления карбонильной группы расходуется 2 м лекулы НАДФН. |
|||
Четвертаяреакциязавершаетодинцикл,в результате которогоприсоединяется |
|||
двухуглеродный фрагментиобразуется радикал бутирила (4 углерода), связанный с |
|||
субъединицей синтазы жирных кислот. Вновь образованная удлиненная ацильная |
|||
группа занимает положение при той SН-группе, с которой исходно была связана |
|||
малонильнаягруппа. |
|
переносится |
из позиции2впозицию 1, |
Перед вторымциклом радикалбутирила |
|
||
(фермент ацилтрансфераза), где находился ацетил в начале первого цикла реакций |
|||
(5). |
|
|
|
Новый цикл реакций, приводящий к удлинению цепи еще на одно двухуглеродное |
|||
звено, начинается с переноса следующей малонильной группы с малонил-СоА на SН- |
|||
группу фосфопантотеина АПБ. |
Затем остаток бутирила подвергается тем же |
||
превращениямиудлиняетсяна2углеродныхатома, происходящихизмалонил-КоА. После таких циклов образуется конечный продукт, пальмитоил –АПБ, который под действиемсеми тиоэстеразы гидролитически отделяется от ферментного комплекса, превращаясь всвободную пальмитиновую кислоту (6).
Синтезированная пальмитиновая кислота при необходимости поступает в эндоплазматический ретикулум или в митохондрии. В Эндоплазматическом Ретикулуме, с участием малонил-КоА и НАДФН цепь удлиняется до С18 или С20. Ферменты, участвующие в элонгации, могут использовать в качестве субстратов не только пальмитиновую, но и другие жирные кислоты, поэтому в организме могут
синтезироваться не только стеариновая кислота, но и жирные кислоты с большим числом атомов углерода. В печени Основной продукт элонгации - стеариновая кислота (С 18:0), однако в ткани мозга образуется большое количество жирных кислот с более длинной цепью - от С20 до С24, которые необходимы для образования сфинголипидов и гликолипидов. Удлиняться могут и ненасыщенные жирные кислоты (олеиновая, линолевая, линоленовая) с образованием производных эйкозановой кислоты (С20).
Для образования двойной связи в радикале жирной кислоты требуется молекулярный кислород, NADH, цитохром b5 и FAD-зависимая редуктаза цитохрома b5. Атомы водорода, отщепляемые от насыщенной кислоты, выделяются в виде воды. Один атом молекулярного кислорода включается в молекулу воды, а другой также восстанавливается до воды с участием электронов NADH, которые передаютсячерез FADH2 ицитохром b5.
Животные организмы могут синтезировать только мононенасыщенные жирные кислоты ––пальмитолеиновую (16 С) и олеиновую (18 С), каждая из которых содержит одну цис-двойную связь. (двойная связь животными клетками вводится не далее 9 атома углерода), поэтому ω-3 и ω-6 полиненасыщенные жирные кислоты не синтезируются в организме, являются незаменимыми и обязательно должны поступатьс пищей,так как выполняютважные регуляторныефункции.
Биосинтез триацилглицеролов(триглицеридов)и фосфолипидов
Триглицериды ифосфолипиды синтезируютсяиз2предшественников: 1.Глицеринфосфата (черездиоксиацетонфосфатизглюкозыилипри фосфорилированиисвободногоглицерола)
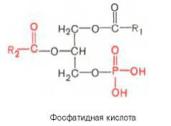
2.ЭфировжирныхкислотсСоА (ацил-СоА) Промежуточным продуктом синтеза является
, которая синтезируется ри связываниифосфатиднаяглицеролкислота-3-фосфата с жирными кислотамип . Фосфатидная кислота можетпревращаться двумя путями:
: фосфатиднаякислотагидролизуется собразованием 1,Синтез2-диацилглицеролатриацилглицер. Последнийлов взаимодействуетс третьеймолекулойСоАпроизводного жирной кислоты, превращается в триацилглицерол. (для того чтобы возниклакаждая эфирнаясвязь,жирнаякислота сначаладолжна активироваться СоА,длячего необходимаэнергияАТФ).
фосфатиднаякислотагидролизуется собразованием 1,Синтез2-диацилглицеролафосфолипидов. Д:иацилглицерол взаимодействует с спиртовым компонентом, который предварительно должен активироваться цитидинтрифосфатом. Фосфорилированные спиртовые группы образуют головы полярных молекул: фосфатидилхолина, фосфатидилсерина, фосфатидилэтаноламина, фосфатидил-
инозитола, которые входят в состав биологических мембран. Фосфолипиды синтезируются в эндоплазматическом ретикулуме, затем переносятся в виде везикул в мембраны других органоидов и встраиваются в них. Таким образом в клеткахсуществует потоквновьсинтезированных полярныхлипидов.
ХолестеролСинтез х естерола— стероид, характерный только для животных организмов. Он синтезируется во многих тканях человека, но основное место синтеза — печень. В печени синтезируется более 50% холестерола, в тонком кишечнике — 15-20%, остальной холестерол синтезируется в коже, коре надпочечников, половых железах. В сутки в организме синтезируется около 1 г холестерола; Холестерол выполняет много функций: входит в состав всех мембран клеток и влияет на их свойства, служит исходным субстратом в . Предшественники в метаболическоминтезепутижесинтезалчныххокислотлестеролаи превращаютсяроидных гормоновтакже вубихинон —компонентдыхательнойцепи.
Холестерол и его эфиры — гидрофобные молекулы, поэтому они транспортируются кровью только в составе разных типов липопротеинов. Нарушения обмена холестерола приводят к одному из наиболее распространённых заболеваний — атеросклерозу. Смертность от последствий атеросклероза (инфаркт миокарда, инсульт) лидирует в общей структуре смертности населения. Накопление холестерола в организме приводит к развитию и другого распространённого заболевания — желчнокаменной болезни. Холестерол синтезируется из ацетил-СоА. Обмен холестерола чрезвычайно сложен — только для его синтеза необходимо осуществление около 100 последовательных реакций. Всего в обмене холестерола участвуетоколо 300 разныхбелков.