146 UNIT 2 ■ Basic Tissues
A
Proerythroblast |
|
Basophilic |
|
Polychromatophilic |
|
Orthochromatophilic |
Erythrocytes |
|
erythroblast |
|
erythroblast |
erythroblast (normoblast) |
|
|
|
|
|
|
|
|
|
Proerythroblast |
Basophilic |
Polychromatophilic |
Orthochromatophilic |
|
|
erythroblast |
Reticulocyte |
Erythrocytes |
|
erythroblast |
erythroblast |
(normoblast) |
|
|
D. Cui
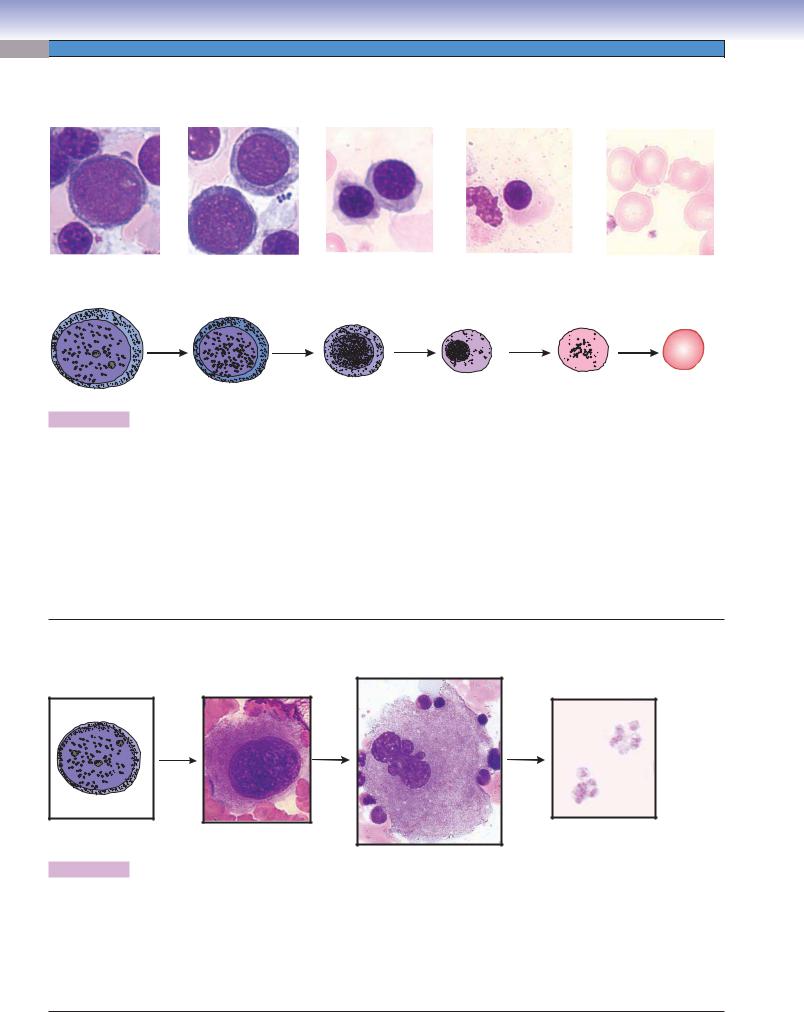
Figure 8-9A. A representation of erythropoiesis (red blood cell formation), bone marrow. Wright stain, 1,569
Erythrocyte formation includes several stages of cell changes during differentiation. Erythrocytes derive from progenitor cells (CFC-Es) that give rise to the first recognizable erythrocyte precursor, the proerythroblast. The proerythroblast is a large cell, which has a large active nucleus with nucleoli. Each proerythroblast divides into two basophilic erythroblasts. Each basophilic erythroblast divides into two polychromatophilic erythroblasts, each of which then divides to form orthochromatophilic erythroblasts, which do not divide. These, in turn, differentiate into reticulocytes (Fig. 8-11B,C), which finally become erythrocytes. There are some general tendencies accompanying differentiation of erythrocytes: (1) the overall size of the cells decreases, (2) the nucleus size decreases and the condensation of the chromatin increases, (3) nucleoli disappear, and (4) the color of the cytoplasm changes from blue to gray to pink because of a reduction of ribosomes and an increase of hemoglobin. When the ribosomes are diluted by cell division and the hemoglobin concentration rises to a near mature level, the cell becomes an orthochromatophilic erythroblast (or normoblast). When the nucleus is extruded and only a few organelles (polyribosomes and mitochondria) remain in the cytoplasm, the cell is called a reticulocyte. The reticulocyte completes maturation and enters the blood circulation to become a mature erythrocyte (red blood corpuscle).
B
Megakaryocyte
Megakaryoblast |
Promegakaryocyte |
Platelets |
D. Cui
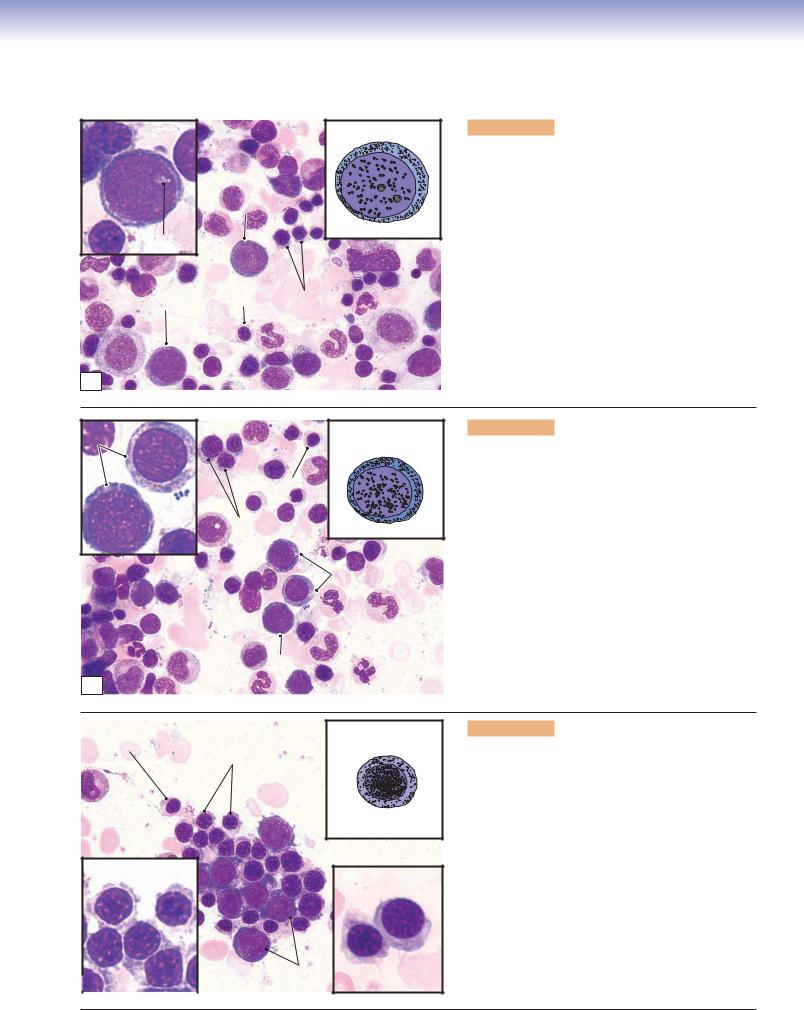
Figure 8-9B. Thrombopoiesis (platelet formation process), bone marrow. Wright stain, 843, 586, and 1,570 (from left to right)
Platelets are very small fragments of cells that have no nuclei. They are also called thrombocytes. Their differentiation from a large cell, the megakaryocyte, takes place in the bone marrow. Megakaryoblasts are the precursor cells. They have a large, round nucleus, undergo mitosis, and become promegakaryocytes. These cells have a large, round nucleus and develop through growth and a series of endomitoses into megakaryocytes. A megakaryocyte has a large, multilobed nucleus with a huge amount of cytoplasm containing numerous granules. The maturation process includes the development of a demarcation membrane system and the subdivision of the cytoplasm to form platelets (Fig. 8-12A,B).

148 UNIT 2 ■ Basic Tissues
|
A |
PoE |
Orthochromatophilic |
Figure 8-11A. Orthochromatophilic erythroblasts, bone |
|
marrow smear. Wright stain, 710; inset 1,569 |
|
|
erythroblast |
|
|
|
|
|
|
|
|
The orthochromatophilic erythroblast, also called a |
|
|
|
|
normoblast, is a very small cell, close to the size of an |
|
PoE |
|
|
erythrocyte. The nucleus is small and so condensed that |
|
|
D. Cui |
it looks like a dark dot because of the extreme condensa- |
|
|
|
|
tion of the chromatin. The cytoplasm appears pinker than |
|
|
|
|
that of the polychromatophilic erythroblast. Hemoglobin |
|
|
|
Orthochromatophilic |
production and accumulation are almost complete, with |
|
|
|
few ribosomes left in the cytoplasm. At this stage, the cell |
|
|
|
erythroblast |
|
|
|
|
is unable to divide. Orthochromatophilic erythroblasts |
|
|
Proerythroblast |
|
become reticulocytes (Fig. 8-11B) after losing their nuclei. |
|
|
|
|
|
OE |
|
|
|
|
B |
|
|
Figure 8-11B. Reticulocytes: the final step of erythro- |
|
Orthochromatophilic |
|
|
cyte formation. |
|
|
|
|
|
erythroblast |
|
Reticulocyte |
|
|
(normoblast) |
|
Orthochromatophilic erythroblasts have small and highly |
|
|
|
|
|
|
|
condensed nuclei. In the next stage, the nucleus is extruded |
|
|
|
|
and phagocytosed by macrophages. Although the cells |
|
|
|
|
loose their nuclei, they retain some polyribosomes in their |
|
|
|
|
cytoplasm. When stained supravitally with cresyl blue or |
|
Mature |
|
|
new methylene blue, the ribosomes aggregate into a blue |
|
|
|
reticular network; therefore, the cells are called reticulo- |
|
erythrocyte |
|
|
cytes. Reticular cells appear the same as mature erythro- |
|
(inside of capillary) |
|
|
cytes with Wright stain (Fig. 8-11C). Reticulocytes enter |
|
|
|
|
|
|
|
|
the blood circulation through the bone marrow sinusoidal |
|
|
|
|
capillaries and become mature erythrocytes in one or two |
|
|
|
|
days. Mature erythrocytes have neither nuclei nor organ- |
|
|
|
|
elles and appear as a biconcave disk. |
|
D. Cui |
|
|
|
|
|
Capillaries |
|
|
CLINICAL CORRELATION
C
Erythrocyte
 Reticulocytes
Reticulocytes
Figure 8-11C. Reticulocytosis, Peripheral Blood Smear. New methylene blue stain, 1,020
Reticulocytosis is a condition characterized by an increased number of reticulocytes. Reticulocytes are premature red blood cells. The normal percentage of reticulocytes is 0.5% to 1.5%. Hemolytic anemia usually increases erythropoietin production, which in turn causes the bone marrow to produce more red blood cells, resulting in a reticulocyte percentage of above 4% to 5%. An increased number of reticulocytes in peripheral blood is an important indication of hemolysis (red blood cell rupture) or bleeding. It can also be the consequence of treating the anemia of chronic kidney disease with erythropoietin. This illustration shows the increased number of reticulocytes with new methylene blue stain after hemolytic anemia.

150 UNIT 2 ■ Basic Tissues
Granulocytopoiesis
A
Myeloblast |
Neutrophilic |
Neutrophilic |
Neutrophilic |
Mature |
|
myelocyte |
metamyelocyte |
stab cell |
neutrophil |
Eosinophilic |
Eosinophilic |
Eosinophilic |
Mature |
myelocyte |
metamyelocyte |
stab cell |
eosinophil |
Promyelocyte
Basophilic |
Basophilic |
Basophilic |
Mature |
myelocyte |
metamyelocyte |
stab cell |
basophil |
D. Cui
Figure 8-13A. A representation of granulocytopoiesis.
In addition to erythropoiesis, leukopoiesis also occurs in the bone marrow. Here are examples of the development of granular leukocytes (granulocytopoiesis). The myeloblast is the earliest morphologically recognizable precursor cell. Cell division occurs in myeloblasts, promyelocytes, and myelocytes. The myelocyte is the last stage that is capable of dividing. The generation of nonspecific granules occurs in the promyelocyte stage and specific granules in the myelocyte stage. Maturation of granulocytes follows this sequence: myeloblasts, promyelocytes, myelocytes, metamyelocytes, and stab (band) cells. The following morphologic changes occur during granulocyte maturation: (1) nucleoli are present only before and during the promyelocyte stage; (2) the nucleus takes the following shapes in different developmental stages: oval, elongated, indented, arched, and then segmented (lobed); and (3) specific granules are first present at the myelocyte stage.
Neutrophilic |
Neutrophilic |
Neutrophilic |
Neutrophil |
myelocyte |
metamyelocyte |
stab cell |
Promyelocyte
Eosinophilic |
Eosinophilic |
Eosinophilic |
Eosinophil |
myelocyte |
metamyelocyte |
stab cell |
|
|
|
|
|
|
|
Figure 8-13B. Overview of stages of granulocytes in development, bone marrow smear. Wright stain, 1,569
These are examples of microphotographs that show various stages of granulocyte maturation in the neutrophilic and eosinophilic series.
152 UNIT 2 ■ Basic Tissues
Neutrophilic |
Neutrophilic Neutrophilic stab cells |
Figure 8-15A. Stab (band) cells, bone marrow smear. Wright |
stab cell |
stab cell |
|
stain, 710; insets 1,569 |
|
|
|
Granular metamyelocytes mature to become stab cells, which |
|
Eosinophilic |
|
are also called band cells. The stab cells (mainly neutrophilic |
|
|
stab cells) can be found in both the bone marrow and the |
|
myelocyte |
|
|
D. Cui |
|
peripheral blood. Their nuclei are elongated and become band |
|
Eosinophilic |
|
and arch (or “C”) shaped, and the cytoplasm is the same as |
Eosinophilic |
metamyelocyte |
|
that of mature neutrophils. These cells are the last stage of |
|
|
granulocyte maturation without division, and in function and |
stab cell |
Basophilic |
|
|
|
structure, they are very close to mature neutrophils. The nuclei |
|
erythroblast |
|
of mature neutrophils become multilobulated (segmented), |
|
|
|
|
|
|
contain dense heterochromatin, and often are described as |
|
|
|
polymorphonuclear (or segmented) neutrophils. |
A |
Neutrophilic |
Neutrophilic |
|
myelocyte |
|
stab cell |
|
Orthochromatophilic |
Polychromatophilic |
Neutrophilic |
erythroblast |
erythroblasts |
|
|
metamyelocyte |
Eosinophil
|
|
Neutrophil |
Neutrophilic |
|
|
|
|
Proerythroblast |
|
stab cell |
B |
|
|
|
|
Figure 8-15B. Bone marrow cells, bone marrow smear. Wright stain, 710
This is an example of blood cells at various stages of development in the bone marrow, which includes both the erythrocyte and the granulocyte series. These cells, at various stages of development, are densely packed together and can be found randomly distributed in the bone marrow. During the maturation process, the cell size becomes smaller and nuclei become denser. In the erythropoiesis series, the cytoplasm of cells becomes light blue and then more pink, and nuclei become much denser and smaller and finally disappear. In the granulocytopoiesis series, the cytoplasm becomes less blue, primary (nonspecific) granules are produced, and then specific granules are produced and are present in myelocytes, which gives these cells the appearance characteristic of their identity as a neutrophil, eosinophil, or basophil. In other changes, nuclei become progressively denser, and the shape changes from round to oval, elongated, indented, and then lobed (segmented).
Blood cells
Reticular tissue
Adipocyte
spaces
C
Figure 8-15C. Bone marrow, bone marrow smear. Wright stain, 35; inset 184
Bone marrow is a specialized example of a reticular connective tissue, a loose connective tissue in which numerous cells are supported by a delicate network of reticular fibers. It resides in cavities within bones (see Figs. 5-8, 5-10, and 5-11). Bone marrow can be categorized into red bone marrow and yellow bone marrow. The term red bone marrow denotes active hematopoiesis; yellow bone marrow refers to a marrow composed chiefly of adipocytes (fat cells). Pictured is a smear of red bone marrow, which contains many developing blood cells, a few adipocytes, and some thin-walled blood vessels (sinusoidal capillaries). The red bone marrow is organized into a hemopoietic compartment and a vascular compartment. The hemopoietic compartment is a network of reticular fibers in which immature and mature blood cells are suspended. The vascular compartment is composed of mainly sinusoidal capillaries, which allow mature blood cells to enter the blood circulation.
CHAPTER 8 ■ Blood and Hemopoiesis |
153 |
Mature neutrophili
or neutrophilic Hematopoietic metamyelocyte compartment
Sinusoidal
capillary
Leukocyte granules
Sinusoidal
capillary
Endothelial cell
 Leukocyte precursors
Leukocyte precursors
Hematopoietic
compartment
Figure 8-16. Developing blood cells in the bone marrow. EM, 5,000
The two compartments of the red (hematopoietically active) bone marrow can be distinguished here. The vascular compartment is composed of sinusoidal capillaries, which in this view contain numerous mature erythrocytes. The hematopoietic compartment is composed of blood cells and the precursors and progenitors of blood cells suspended in a loose network of support cells and reticular fibers. The cells seen in the hematopoietic compartment here appear to be mostly developing or mature granulocytes. The exact stage of differentiation of individual cells is not as clear here as it is in a bone marrow smear.
154 UNIT 2 ■ Basic Tissues
SYNOPSIS 8 - 2 Hematopoiesis
Stem, Progenitor, and Precursor cells
■Stem cells are capable of differentiating into multiple cell lineages and can undergo proliferation indefinitely.
■Progenitor cells are only capable of differentiating into a single cell lineage (restricted to one or two blood cell types) and are morphologically undifferentiated.
■Precursor cells can be recognized morphologically as undergoing differentiation along a particular blood cell lineage.
Erythropoiesis
■Cytoplasm becomes progressively less basophilic because of dilution of ribosomes during the erythropoiesis process.
■Nucleus size progressively decreases because of increased condensation of chromatin.
■Cell size progressively decreases during the erythroid differentiation.
■Cytoplasm becomes progressively more eosinophilic because of increased accumulation of hemoglobin.
■The nucleus retains a round shape and no indentation occurs before it disappears.
Granulocytopoiesis
■Cell size decreases and nucleus becomes more condensed as in erythropoiesis.
■Nucleus shape changes from round or oval (promyeloblasts) to kidney shaped/slightly indented (myelocytes) and then changes from deeply indented (metamyelocytes) to band shaped (band cells) and finally to lobed (mature granulocytes).
■Promyelocytes do not have specific granules (only azurophilic granules); at this stage, it is too early to tell which granular leukocytes they will become.
■Myelocytes are the last developmental stage capable of dividing; specific granules accumulate in this stage.
SYNOPSIS 8 - 3 Pathological and Clinical Terms for Mature and Developing Blood Cells
■Gray platelet syndrome: This condition is characterized by a deficiency or absence of the alpha granules and contents in blood platelets, giving platelets a gray appearance in a Wright stain smear (Fig. 8-2B).
■Platelet storage pool deficiency: Disorder caused by a decrease or absence of platelet delta granules (dense bodies), which normally store and release adenine nucleotides and 5HT. “Platelet-type” bleeding is common with this deficiency (Fig. 8-2B).
■Petechiae: Minute red or purple spots on the skin or mucous membranes caused by capillary hemorrhage; common causes include physical trauma and decreased platelets (thrombocytopenia).
■Smudge cell: Damaged lymphocytes seen on a peripheral blood smear caused by mechanical stress in the process of producing the smear; although nonspecific, smudge cells are encountered more frequently on blood smears of patients with chronic lymphocytic leukemia (Fig. 8-4C).
■Reticulocytosis: Increased reticulocytes in the blood, often in response to blood loss, stimulation by erythropoietin treatment, or treatment of iron deficiency anemia with iron supplementation (Fig. 8-11C).
■Thrombocytosis: Increased platelet count in the blood, which may be reactive or neoplastic, as in the disease essential thrombocytosis (Fig. 8-12C).

9 Circulatory System
Introduction and Key Concepts for the Circulatory System
The Cardiovascular System
Figure 9-1 |
Overview of the Cardiovascular System |
Figure 9-2 |
The Heart and Its Impulse Conductive Function |
Synopsis 9-1 |
Structure and Functions of the Heart |
Figure 9-3A |
Layers of the Heart Wall: Endocardium, Ventricle |
Figure 9-3B |
Layers of the Heart Wall: Myocardium, Ventricle |
Figure 9-3C |
Layers of the Heart Wall: Epicardium, Ventricle |
Figure 9-4A |
Purkinje Fibers and Intercalated Disks |
Figure 9-4B |
Cardiac Valves |
Figure 9-4C |
Clinical Correlation: Myocardial Infarction |
Figure 9-5 |
A Representation of the General Structure of Blood Vessel Layers (Tunicae) and a |
|
Comparison of the Medium Artery and the Medium Vein |
Figure 9-6 |
A Representation of Types of Blood Vessels: Arteries, Veins, and Capillaries |
The Arterial System |
|
Figure 9-7A–C |
Large Arteries (Elastic Arteries) |
Figure 9-8A–C |
Medium Arteries (Muscular Arteries) |
Figure 9-9A |
Medium Artery |
Figure 9-9B |
Small Artery |
Figure 9-10A–C |
Small Arteries and Arterioles |
Figure 9-11 |
Arteriole |
Synopsis 9-2 |
Functions of Endothelium in Blood Vessels |
Figure 9-12A |
Clinical Correlation: Coronary Artery Atherosclerosis |
Figure 9-12B |
Clinical Correlation: Polyarteritis Nodosa (Vasculitis) |
Synopsis 9-3 |
Pathological and Clinical Terms for the Circulatory System |



 Megakaryocyte
Megakaryocyte


